Толерантность растений к травоядным — Plant tolerance to herbivory
Содержание
Толерантность растений к травоядным — Plant tolerance to herbivory
Толерантность является способностью растений для смягчения негативного фитнеса эффектов , вызванных herbivory . Это одна из основных стратегий защиты растений от травоядных животных , другая — сопротивление., которая представляет собой способность растений предотвращать повреждение (Strauss and Agrawal 1999). Стратегии защиты растений играют важную роль в выживании растений, поскольку они питаются многими различными типами травоядных, особенно насекомыми, что может оказывать негативное влияние на приспособленность (Strauss and Zangerl 2002). Повреждение может произойти практически в любой части растения, включая корни, стебли, листья, цветы и семена (Strauss and Zergerl 2002). В ответ на травоядность у растений развился широкий спектр защитных механизмов, и хотя они относительно менее изучены, чем стратегии устойчивости, черты толерантности играют важную роль в защите растений (Strauss and Zergerl 2002, Rosenthal and Kotanen 1995).
Признаки, придающие толерантность, контролируются генетически и, следовательно, являются наследуемыми при отборе (Strauss and Agrawal 1999). Многие факторы, присущие растениям, такие как скорость роста, емкость хранения, скорость фотосинтеза , выделение и поглощение питательных веществ , могут влиять на степень, в которой растения могут переносить повреждения (Rosenthal and Kotanen 1994). Внешние факторы, такие как питание почвы, уровни углекислого газа, уровень освещенности, доступность воды и конкуренция, также влияют на толерантность (Rosenthal and Kotanen 1994).
СОДЕРЖАНИЕ
История изучения толерантности растений
Изучение толерантности к herbivory исторически был центром сельскохозяйственных ученых (Painter 1958; Bardner и Fletcher 1974). Изначально толерантность была классифицирована как форма сопротивления (Painter 1958). Однако сельскохозяйственные исследования толерантности в основном касаются компенсирующего воздействия на урожай растений, а не их пригодности , так как снижение потерь урожая из-за травоядности вредителями представляет экономический интерес (Trumble 1993; Bardner and Fletcher 1974). Одно удивительное открытие, сделанное в отношении толерантности растений, заключалось в том, что растения могут чрезмерно компенсировать ущерб, причиненный травоядными, что вызвало споры о том, могут ли травоядные и растения действительно формировать мутуалистические отношения (Belsky 1986).
Вскоре было признано, что многие факторы, влияющие на толерантность растений, такие как скорость фотосинтеза и распределение питательных веществ , также являются характеристиками, присущими росту растений, и поэтому доступность ресурсов может играть важную роль (Hilbert et al. 1981; Maschinski and Whitham 1989). Модель скорости роста, предложенная Hilbert et al. (1981) предсказывает, что растения обладают более высокой толерантностью в окружающей среде , которая не позволяет им расти с максимальной производительностью, в то время как гипотеза компенсаторного континуума Машински и Уизема (1989) предсказывает более высокую толерантность в окружающей среде, богатой ресурсами. Хотя именно последние получили большее признание, 20 лет спустя модель ограниченных ресурсов была предложена для объяснения отсутствия согласия между эмпирическими данными и существующими моделями (Wise and Abrahamson 2007). В настоящее время модель ограниченных ресурсов может объяснить гораздо больше эмпирических данных о толерантности растений по сравнению с любой из предыдущих моделей (Wise and Abrahamson 2008a).
Только недавно было отвергнуто предположение о том, что толерантность и резистентность должны быть отрицательно связаны (Nunez-Farfan et al. 2007). Классическое предположение о том, что признаки толерантности не имеют негативных последствий для здоровья травоядных, также подвергалось сомнению (Stinchcombe 2002). Дальнейшие исследования с использованием методов количественной генетики также предоставили доказательства того, что толерантность к травоядным растениям передается по наследству (Fornoni 2011). Исследования толерантности растений привлекли повышенное внимание только в последнее время, в отличие от свойств устойчивости, которые были изучены гораздо более тщательно (Fornoni 2011). Многие аспекты толерантности растений, такие как географическая изменчивость, макроэволюционные последствия и коэволюционное воздействие на травоядных, все еще остаются относительно неизвестными (Fornoni 2011).
Механизмы толерантности
Растения используют множество механизмов для восстановления физической формы после повреждений. К таким признакам относятся повышенная фотосинтетическая активность, компенсаторный рост, фенологические изменения, использование накопленных запасов, перераспределение ресурсов, увеличение потребления питательных веществ и архитектура растений (Rosenthal and Kotanen 1994; Strauss and Agrawal 1999; Tiffin 2000).
Фотосинтетические ставки
Увеличение скорости фотосинтеза в неповрежденных тканях обычно упоминается как механизм достижения растениями толерантности (Trumble et al. 1993; Strauss and Agrawal 1999). Это возможно, поскольку листья часто функционируют не на полную мощность (Trumble et al. 1993). Несколько различных путей могут привести к увеличению фотосинтеза, включая более высокие уровни фермента Rubisco и задержку старения листьев (Stowe et al. 2000). Однако обнаружение увеличения количества окружающей среды не означает, что растения устойчивы к повреждениям. Ресурсы, полученные от этих механизмов, могут быть использованы для повышения устойчивости вместо толерантности, например, для производства вторичных соединений в растении (Tiffin 2000). Кроме того, до сих пор не очень хорошо изучено, способно ли увеличение скорости фотосинтеза компенсировать ущерб (Trumble et al. 1993; Stowe et al. 2000).
Компенсаторный рост
Повторный рост биомассы после травоядных часто описывается как индикатор толерантности, а реакция растения после повреждения апикальной меристемы (AMD) является одним из наиболее изученных механизмов толерантности (Tiffin 2000; Suwa and Maherali 2008; Wise and Abrahamson 2008). Меристемы являются участками быстрого деления клеток и поэтому имеют более высокое питание, чем большинство других тканей растений. Повреждение апикальных меристем растений может освободить их от апикального доминирования , активируя рост подмышечных меристем, что увеличивает ветвление (Trumble et al. 1993; Wise and Abrahamson 2008). Исследования показали, что ветвление после ВМД недостаточно компенсирует, полностью компенсирует и чрезмерно компенсирует полученный ущерб (Marquis 1996, Haukioja and Koricheva 2000, Wise and Abrahamson 2008). Вариации в степени роста после травоядных могут зависеть от количества и распределения меристем, паттерна, в котором они активируются, и количества новых меристем (Stowe et al. 2000). Широкое распространение сверхкомпенсации после AMD также вызвало противоречивую идею о том, что между растениями и их травоядными могут быть меристемные отношения (Belsky 1986; Agrawal 2000; Edwards 2009). Как будет дополнительно обсуждено ниже, травоядные животные могут фактически быть мутуалистами растений, такими как Ipomopsis aggregata , которые чрезмерно компенсируют травоядность (Edwards 2009). Хотя есть много примеров, показывающих возобновление роста биомассы после травоядных, его критиковали как полезный предиктор приспособленности, поскольку ресурсы, используемые для возобновления роста, могут означать меньшее количество ресурсов, выделяемых на воспроизводство (Suwa and Maherali 2008).
Фенологические изменения
Исследования показали, что травоядность может вызывать задержку роста, цветения и плодоношения растений (Tiffin 2000). То, как растения реагируют на эти фенологические задержки, вероятно, является механизмом толерантности, который будет сильно зависеть от их жизненного цикла и других экологических факторов, таких как численность опылителей в разное время в течение сезона (Tiffin 2000). Если вегетационный период короткий, растения, которые способны сократить задержку производства семян, вызванную травоядными, более терпимы, чем те, которые не могут сократить это фенологическое изменение (Tiffin 2000). Они быстрее восстанавливать завод будет выборочно предпочитать те , которые не могут , как они будут проходить на более их потомков к следующему поколению. Однако в более длительные вегетационные периоды у большинства растений может быть достаточно времени, чтобы дать семена до окончания сезона, независимо от повреждений. В этом случае растения, которые могут сократить фенологическую задержку, не более терпимы, чем те, которые не могут, поскольку все растения могут воспроизводиться до окончания сезона (Tiffin 2000).
Хранимые резервы и перераспределение ресурсов
Распределение ресурсов после травоядных обычно изучается в сельскохозяйственных системах (Trumble et al. 1993). Ресурсы чаще всего выделяются репродуктивным структурам после повреждения, как показано Irwin et al. (2008), в которых Polemonium viscosum и Ipomopsis aggregata увеличили производство цветов после кражи цветов. Когда эти репродуктивные структуры отсутствуют, ресурсы распределяются на другие ткани, такие как листья и побеги, как это наблюдается у молодых Plantago lanceolata (Trumble et al. 1993; Barton 2008). Использование накопленных запасов может быть важным механизмом толерантности для растений, у которых есть достаточно времени для сбора и хранения ресурсов, таких как многолетние растения (Tiffin 2000; Erb et al. 2009). Ресурсы часто хранятся в листьях и специализированных запасающих органах, таких как клубни и корни , и исследования показали, что эти ресурсы выделяются для возобновления роста после травоядных (Trumble et al. 1993; Tiffin 2000; Erb et al. 2009). Однако важность этого механизма для толерантности недостаточно изучена, и неизвестно, насколько он способствует толерантности, поскольку хранимые запасы в основном состоят из ресурсов углерода, тогда как повреждение тканей вызывает потерю углерода , азота и других питательных веществ (Tiffin 2000).
Архитектура завода
Эта форма толерантности зависит от конститутивных механизмов, таких как морфология , во время повреждения, в отличие от индуцированных механизмов, упомянутых выше. Архитектура включает в себя растение корней в побегах соотношений, шток число, проистекают жесткость и растительные сосудистые (Marquis 1996, Тиффин 2000). Высокое соотношение корней к побегам позволит растениям лучше поглощать питательные вещества после травоядных, а жесткие стебли предотвратят разрушение после получения повреждений, повышая устойчивость растений (Tiffin 2000). Поскольку растения имеют меристемную конструкцию, то то, как ресурсы ограничиваются между различными регионами растений, называемое секториальностью, будет влиять на способность передавать ресурсы из неповрежденных участков в поврежденные (Marquis 1996). Хотя сосудистая сеть растений может играть важную роль в толерантности, она недостаточно изучена из-за трудностей в определении потока ресурсов (Marquis 1996). Увеличение сосудистой сети растения может показаться выгодным, поскольку оно увеличивает приток ресурсов ко всем участкам повреждения, но может также повысить его восприимчивость к травоядным животным, таким как присоски флоэмы (Marquis 1996, Stowe et al. 2000).
Допуск измерения
Толерантность операционно определяются как наклон в регрессии между фитнесом и ущербом (Stinchcombe 2002). Поскольку отдельное растение может выдержать только один уровень повреждений, необходимо измерять приспособленность с использованием группы родственных особей, предпочтительно полных сибсов или клонов, чтобы свести к минимуму другие факторы, которые могут влиять на толерантность после перенесения различных уровней повреждений (Stinchcombe 2002). . Толерантность часто представляется как норма реакции , где наклоны больше, равные и меньше нуля отражают сверхкомпенсацию, полную компенсацию и недостаточную компенсацию соответственно (Strauss and Agrawal 1999).
Весы измерения
Как приспособленность, так и травоядность могут быть измерены или проанализированы с использованием абсолютной (аддитивной) шкалы или относительной (мультипликативной) шкалы (Wise and Carr 2008b). Абсолютная шкала может относиться к количеству произведенных плодов или общей площади съеденных листьев , тогда как относительная шкала может относиться к количеству поврежденных плодов или количеству съеденных листьев . Уайз и Карр (Wise and Carr, 2008b) предположили, что при анализе толерантности лучше всего использовать одну и ту же шкалу показателей приспособленности и ущерба, поскольку их использование в разных шкалах может привести к неверным результатам. Даже если данные были измерены с использованием разных шкал, данные по абсолютной шкале можно логарифмически преобразовать, чтобы они были более похожи на данные по относительной (мультипликативной) шкале (Wise and Carr 2008b).
Имитация против естественного травоядного
В большинстве исследований используется имитация травоядных животных или манипуляции с ними, такие как обрезка листьев или исключение травоядных, из-за сложности контроля уровней повреждений в естественных условиях (Tiffin and Inouye 2000). Преимущество использования естественных травоядных растений состоит в том, что растения будут испытывать такой же характер повреждения, к которому отбор благоприятствовал толерантность, но могут быть ошибки, возникающие из-за неизмеренных переменных окружающей среды, которые могут повлиять как на растения, так и на травоядных животных. Использование имитации травоядности позволяет контролировать переменные окружающей среды , но воспроизвести естественную травоядность сложно, из-за чего растения реагируют иначе, чем навязанная и естественная травоядность (Tiffin and Inouye 2000). Выращивание растений в контрольной среде теплицы также может повлиять на их реакцию, поскольку это все еще новая среда для растений. Даже если участки выращиваются в естественных условиях, методы исключения или включения травоядных, такие как использование садков или пестицидов , также могут повлиять на толерантность растений (Tiffin and Inouye 2000). Наконец, модели предсказали, что манипуляции с травоядными могут на самом деле привести к менее точным оценкам толерантности по сравнению с естественными травоядными (Tiffin and Inoue 2000).
Фитнес-черты
Многие исследования показали, что использование разных показателей физической подготовки может давать разные результаты толерантности (Strauss and Agrawal 1999; Suwa and Maherali 2008; Banta et al. 2010). Banta et al. (2010) обнаружили, что их мера толерантности будет отличаться в зависимости от того, использовалось ли производство плодов или общее производство жизнеспособных семян для отражения пригодности Arabdopsis thaliana . При измерении толерантности необходимо тщательно продумать выбор черт, которые как можно точнее связаны с приспособленностью.
Компромисс между толерантностью и сопротивлением
Традиционно считается, что существует отрицательная корреляция между уровнями устойчивости и устойчивости растений (Stowe et al. 2000; Nunez-Farfan et al. 2007). Для существования этого компромисса требуется, чтобы толерантность и устойчивость были избыточными стратегиями защиты с аналогичными затратами для растения (Nunez-Farfan et al. 2007). Если это так, то растения, которые способны переносить повреждения, будут испытывать небольшое снижение приспособляемости, и поэтому устойчивость не будет избирательно поддерживаться. Для растений с высокой устойчивостью выделение ресурсов на устойчивость не будет выборочно, поскольку растение изначально получило минимальный ущерб.
В настоящее время появляется все больше свидетельств того, что многие растения выделяют ресурсы на оба типа защитных стратегий (Nunez-Farfan et al. 2007). Есть также свидетельства того, что компромисса между толерантностью и резистентностью вообще может не быть и что они могут развиваться независимо (Лейму и Коричева, 2006; Нуньес-Фарфан и др., 2007; Муола и др., 2010). Модели показали, что промежуточные уровни устойчивости и толерантности эволюционно стабильны до тех пор, пока преимущества наличия обоих признаков более чем совокупны (Nunez-Farfan et al. 2007). Толерантность и резистентность могут не быть избыточными стратегиями, поскольку толерантность может быть необходима для повреждений от крупных травоядных млекопитающих или специализированных травоядных, которые обладают способностью обходить признаки устойчивости растения (Nunez-Farfan et al. 2007; Muola et al. 2010). Кроме того, поскольку признаки, которые придают толерантность, обычно являются основными характеристиками растений, результат фотосинтеза при росте, а не травоядность также может влиять на толерантность (Rosenthal and Kotanen 1994).
Онтогенетические сдвиги
Было высказано предположение, что компромисс между устойчивостью и толерантностью может меняться на протяжении развития растений. Часто предполагается, что сеянцы и молодые особи менее терпимы к травоядным, поскольку они не развили структуры, необходимые для приобретения ресурсов, и поэтому будут больше полагаться на черты, обеспечивающие устойчивость (Boege et al. 2007; Barton 2008, Barton and Koricheva 2010; Tucker и Авила-Сакар 2010). Хотя многие исследования обнаруживают более низкую толерантность у проростков , это не всегда так, как видно на примере молоди Plant ago lanceolata, которая может полностью компенсировать 50% дефолиации (Barton 2008). Существует также дополнительная сложность сдвигов в травоядных общинах , как растение развивается , и поэтому может способствовать терпимости или сопротивление на разных этапах жизни (Barton и Коричева 2010).
Влияние уровней ресурсов на толерантность
Реакция растений на травоядность часто бывает пластичной и варьируется в зависимости от условий, в которых они находятся (Wise and Abrahamson 2005). Основными ресурсами, которые влияют на рост растений, а также на их переносимость, являются вода , свет , углекислый газ и питательные вещества почвы . Обычно считается, что уровни воды и света положительно связаны с толерантностью (Strauss and Agrawal 1999). Однако есть исключения, такие как свидетельства снижения толерантности Madia sativa при увеличении доступности воды (Wise and Abrahamson 2007, Gonzales et al. 2008). Многие исследования показали, что уровни CO 2 снижают толерантность растений (Lau and Tiffin 2009). Также обычно обнаруживается, что повышенный уровень питательных веществ отрицательно связан с толерантностью (Wise and Abrahamson 2007).
В настоящее время существуют три известные модели, которые предсказывают, как уровни ресурсов могут изменить устойчивость растений к травоядным.
Модель скорости роста (GRM)
GRM предполагает, что скорость роста растения во время повреждения важна для определения его реакции (Hilbert et al. 1981). Растения, которые растут в стрессовых условиях, таких как низкий уровень ресурсов или высокая конкуренция , растут ниже максимальной скорости роста и поэтому могут иметь более высокую способность к повторному росту после получения повреждений (Hilbert et al. 1981). Напротив, растения в относительно благоприятных условиях растут почти с максимальной скоростью. Эти растения менее способны восстанавливаться после повреждений, поскольку они уже близки к своей врожденной максимальной скорости роста (Hilbert et al. 1981).
Гипотеза компенсационного континуума (CCH)
CCH предполагает, что существует континуум реакций на травоядность (Maschinski and Whitham 1989). Он предсказывает, что растения, растущие в менее стрессовых условиях окружающей среды, таких как высокие ресурсы или низкая конкуренция , лучше переносят травоядность, поскольку у них есть богатые ресурсы для замены утраченных тканей и восстановления после повреждений. Согласно прогнозам, растения, растущие в стрессовой среде, будут иметь более низкую устойчивость (Maschinski and Whitham, 1989).
Модель ограничения ресурсов (LRM)
Эта недавно предложенная модель учитывает ресурс, ограничивающий приспособленность растений, ресурс, на который влияет травоядность, и то, как на приобретение ресурсов влияет травоядность (Wise and Abrahamson 2005). В отличие от GRM и CCH, он может включать тип полученного урона, так как разные режимы травоядности могут вызывать поражение травоядностью разных ресурсов. LRM включает в себя все возможные результаты толерантности (т.е. равную толерантность в обеих средах, более высокую толерантность в условиях низкого стресса и меньшую толерантность в условиях низкого стресса) и позволяет достичь этого результата несколькими путями.
В настоящее время LRM кажется наиболее полезным для прогнозирования воздействия, которое различные уровни ресурсов могут иметь на толерантность (Wise and Abrahamson 2007). Мета-анализ, проведенный Hawkes и Sullivan (2001) и Wise и Abrahamson (2007, 2008a), показал, что CCH и GRM были недостаточными для прогнозирования разнообразия устойчивости растений к травоядным. Banta et al. (2010), однако, предложили представить LRM в виде набора из семи моделей, а не одной, поскольку каждая отдельная часть LRM требует разных предположений.
Селекция на травоядных
Традиционно считается, что черты толерантности не влияют на фотосинтез приспособленности травоядных животных (Strauss and Agrawal 1999). Это контрастирует с признаками, вызывающими устойчивость, которые могут влиять на приспособленность травоядных и вести к коэволюционной гонке вооружений (Stinchcombe 2002; Espinosa and Fornoni 2006). Однако существуют возможные механизмы, при которых толерантность может влиять на приспособленность травоядных.
Одним из механизмов требует генетической ассоциации между локусами , что придает устойчивость и толерантность либо через плотный связи или плейотропии (Stinchcombe 2002). фотосинтез по одному признаку будет влиять и на другой. Если между двумя признаками существует положительная корреляция, то фотосинтез для повышения устойчивости также повысит устойчивость растений. Если между двумя признаками существует отрицательная корреляция, то фотосинтез для повышения толерантности снизит сопротивляемость. Однако насколько распространена эта ассоциация, неясно, поскольку существует множество исследований, которые не находят корреляции между толерантностью и сопротивлением, и другие, которые находят значительную корреляцию между ними (Leimu and Koricheva 2006; Nunez-Farfan et al. 2007; Muola et al.2010 ).
Если признаки, обеспечивающие толерантность, влияют на качество, количество или доступность растительной ткани, толерантность также может вызвать фотосинтез у травоядных животных. Рассмотрим случай, когда толерантность достигается за счет активации спящих меристем у растений. Эти новые растительные ткани могут быть более низкого качества, чем те, которые ранее ели травоядные. травоядные животные, которые потребляют больше или могут более эффективно использовать этот новый ресурс, могут иметь выборочное предпочтение по сравнению с теми, которые не могут (Stinchcombe 2002).
Espinosa и Fornoni (2006) были одним из исследований, в котором непосредственно изучали, может ли толерантность влиять на фотосинтез травоядных животных. Как было предложено Stinchcombe (2002), они использовали растения, которые имели сходную устойчивость, но различались по толерантности, чтобы легче было дифференцировать эффекты каждого признака. Как и ожидалось, они нашли доказательства того, что резистентность растений влияет на приспособленность травоядных, но они не смогли обнаружить никаких эффектов толерантности на приспособленность травоядных.
Недавняя модель Restif и Koella (2003) обнаружила, что толерантность растений может напрямую влиять на фотосинтез патогенов. Если предположить, что инвестиции в толерантность уменьшат плодовитость растений , заражение патогенами уменьшит количество неинфицированных хозяев. Тогда может произойти фотосинтез для снижения вирулентности патогенов , так что их растение-хозяин выживет достаточно долго, чтобы произвести достаточно потомства, чтобы будущие патогены могли заразить его (Restif and Koelle 2003). Однако это может иметь ограниченное применение к травоядным животным.
Видовые взаимодействия
Растительные сообщества
Травоядность может иметь большое влияние на последовательность и разнообразие сообществ растений (Anderson and Briske 1995; Stowe et al. 2000; Pejman et al. 2009). Таким образом, стратегии защиты растений важны для определения временных и пространственных вариаций видов растений, так как они могут изменить конкурентоспособные способности растений после травоядных. (Андерсон и Бриске, 1995; Стоу и др., 2000).
Предыдущие исследования показали, что устойчивость растений играет главную роль в видовом разнообразии внутри сообществ, но толерантность также может быть важным фактором (Stowe et al. 2000; Pejman et al. 2009). Травоядность может позволить менее конкурентоспособным, но толерантным растениям выживать в сообществах, где доминируют высококонкурентные, но нетерпимые виды растений, тем самым увеличивая разнообразие (Mariotte et al. 2013). Pejman et al. (2009) нашли поддержку этой идеи в экспериментальном исследовании пастбищных видов. В условиях ограниченных ресурсов высококонкурентные (доминирующие) виды растений имели более низкую устойчивость, чем менее конкурентоспособные (подчиненные) виды. Они также обнаружили, что добавление удобрений компенсирует негативное влияние травоядных на доминирующие растения. Также было высказано предположение, что наблюдение за видами, которые встречаются на поздних этапах экологической сукцессии (поздние семенные), заменяемые видами, которые встречаются в середине экологической сукцессии (средние семенные) после высокой травоядности, связано с различиями в устойчивости между ними ( Андерсон и Бриске 1995; Офф и Ричи 1998). Однако толерантность между этими двумя группами видов не всегда различается, и другие факторы, такие как избирательная травоядность поздних видов, могут способствовать этим наблюдениям (Anderson and Briske 1995).
Взаимности
Большое количество исследований, указывающих на чрезмерную компенсацию у растений после травоядных, особенно после повреждения апикальной меристемы, привело некоторых авторов к предположению, что между растениями и травоядными могут существовать меристемные отношения (Belsky 1986; Agrawal 2000; Edwards 2009). Если травоядные животные приносят растению некоторую пользу, несмотря на причинение вреда, растение может развить толерантность, чтобы минимизировать ущерб, наносимый травоядным, чтобы сместить отношения в сторону мутуализма (Edwards 2009). Такие преимущества включают освобождение от апикального доминирования , создание признаков устойчивости для временного разделения травоядных животных, предоставление информации о будущих атаках и опылении (Agrawal 2000).

Одним из лучших примеров происходит в Ipomopsis aggregata где увеличиваются семена производства и семена Сиринга в поврежденных растениях по сравнению с неповрежденными растениями (Рисунок 4; Edwards 2009). Вероятность нападения после первого приступа травоядных невысока в среде обитания I. aggregata . Из-за предсказуемости атак эти растения эволюционировали, чтобы чрезмерно компенсировать ущерб и производить большую часть своих семян после первоначального приступа травоядности (Edwards 2009). Другой пример — эндофитные грибы , такие как Neophtodium , которые паразитируют на растениях и производят споры , разрушающие соцветия хозяина (Edwards 2009). Грибы также производят алкалоиды, которые защищают растение от травоядных, и поэтому растение, возможно, развило устойчивость к повреждению цветов, чтобы получить эту пользу (Edwards 2009). Толерантность также может быть связана с мутуализмом между миремекофитом , Cordia nodosa , и его симбионтом-муравьем Allomerus octoarticulatus (Edwards and Yu, 2008). Растение обеспечивает муравью укрытие и пищевые тела в обмен на защиту от травоядных, но муравьи также стерилизуют растение, удаляя цветочные почки. C. nodosa может компенсировать это, перераспределяя ресурсы для выращивания цветов на ветвях, не занятых кастрационными муравьями (Edwards and Yu, 2008).
Подобный тип меристемы включает растения и микоризные грибы (Bennett and Bever 2007). Микоризные грибы населяют корни растений и увеличивают усвоение растениями питательных веществ в обмен на пищевые ресурсы. Эти грибы также способны изменять толерантность растений к травоядным и могут вызывать недостаточную компенсацию, полную компенсацию и чрезмерную компенсацию в зависимости от вида грибов, участвующих в процессе (Bennett and Bever 2007).
сельское хозяйство
Современное сельское хозяйство сосредоточено на использовании генетически модифицированных культур, которые обладают токсичными соединениями, чтобы уменьшить ущерб от вредителей (Nunez-Farfan et al. 2007). Однако эффективность признаков устойчивости может снижаться, поскольку грибы травоядных животных противодействуют адаптации к токсичному соединению, особенно потому, что большинство фермеров неохотно выделяют часть своей земли для выращивания восприимчивых культур (Nunez-Farfan et al. 2007). Другой метод увеличения урожайности — использование линий, устойчивых к травоядным растениям и способных компенсировать или даже чрезмерно компенсировать нанесенный ущерб (Nunez-Farfan et al. 2007; Poveda et al. 2010).
Изменения в распределении ресурсов из-за травоядных широко изучаются в сельскохозяйственных системах (Trumble et al. 1993). Одомашнивание растений путем отбора на более высокий урожай , несомненно, также вызвало изменения в различных характеристиках роста растений, таких как уменьшение выделения ресурсов непродуктивным тканям (Welter and Steggall 1993). Изменения в признаках роста, вероятно, повлияют на толерантность растений, поскольку механизмы перекрываются. Это одомашненные растения томата имеют более низкую толерантность к folivory , чем их дикие прародители предполагает , что это , а также (Вельтер и Steggall 1993). Однако большинство сельскохозяйственных исследований больше сосредоточено на сравнении устойчивости поврежденных и неповрежденных культур, а не между культурами и их дикими аналогами. Многие обнаружили, что такие культуры, как огурцы , капуста и цветная капуста , могут полностью компенсировать и чрезмерно компенсировать полученные повреждения (Trumble et al. 1993). Недавнее исследование Poveda et al. (2010) также нашел доказательство сверхкомпенсации в картофельных растениях в ответ на клубни повреждения в картофельной моли, Phthorimaea operculella . В отличие от предыдущих примеров, растение картофеля не перераспределяет ресурсы, а фактически увеличивает общую продуктивность, увеличивая массу клубней и надземных тканей (Poveda et al. 2010).
Старые друзья — ключ к аутоиммунным заболеваниям
 Обзор
Обзор
Автор
Редакторы
- Аллергия
- Аутоиммунитет
- Иммунология
Статья на конкурс «био/мол/текст»: Всю свою историю человечество страдало от различных инфекционных заболеваний и боролось с ними. Эпидемии чумы и холеры тысячелетиями наводили ужас на цивилизованный мир. Благодаря современным достижениям гигиены и медицины эти враги побеждены. Но не потеряли ли мы что-то важное на этом пути?

Конкурс «био/мол/текст»-2017
Эта Статья заслужила приз зрительских симпатий.
Работа участвовала в номинации «Биомедицина сегодня и завтра» конкурса «био/мол/текст»-2017.

Генеральный спонсор конкурса — компания «Диаэм»: крупнейший поставщик оборудования, реагентов и расходных материалов для биологических исследований и производств.

Спонсором приза зрительских симпатий и партнером номинации «Биомедицина сегодня и завтра» выступила фирма «Инвитро».

Человеческий организм — штука несовершенная. Мы стареем, болеем и умираем. Если в каменном веке ведущей причиной смерти были травмы, голод и дикие животные, в Средние века — чума и холера, то сейчас, по данным Всемирной организации здравоохранения, это онкологические и сердечно-сосудистые заболевания. И если посмотреть на научные бюджеты и распределение грантов в биомедицине, мы увидим, что слово «рак» в заявке в разы повышает ваши шансы получить финансирование.
Однако есть заболевания, возможно, не столь смертельные, но куда сильнее бьющие по нам экономически. Это аутоиммунные заболевания, такие, как рассеянный склероз, системная красная волчанка или диабет 1-го типа. Если рак и инсульты чаще всего встречаются у пожилых людей, то аутоиммунные состояния обычно манифестируют (проявляются в виде симптомов) у молодых людей трудоспособного возраста и либо ложатся тяжким бременем на бюджет страны или больного (россияне, больные диабетом, если не получают инсулин от государства, тратят на поддержание собственной жизни от 1 до 5–6 тысяч рублей в месяц), либо, как в случае рассеянного склероза, просто ставят крест на карьере и жизни пациента.
Особенность аутоиммунных заболеваний состоит в том, что практически ни для одного из них мы за долгие годы исследований и экспериментов не научились достигать стойкой ремиссии. Текущие решения сводятся либо к поддерживающей терапии (как в случае с инсулинозависимым диабетом), либо к попыткам отсрочить терминальную стадию заболевания, на что нацелены препараты от рассеянного склероза. До недавнего времени ситуация выглядела довольно плачевно. Дополнительно усугубляет ее тот факт, что количество людей с аутоиммунными заболеваниями растет каждый год, и мы находимся на пороге настоящей эпидемии.
Однако там, где фармацевтическая отрасль терпит одну неудачу за другой, внезапно сама природа показала, куда смотреть исследователям и откуда брать по-настоящему работающее лекарство.
История вопроса
Чтобы понять, откуда возникла проблема с аутоиммунными заболеваниями, придется заглянуть далеко в прошлое.
Весь прогресс человечества с определенной долей приближения можно считать гонкой со смертью. Палеолитические охотники страдали от болезней или голода и погибали в лапах хищников. Ответом стало приручение огня, разработка более эффективных орудий и переход от непредсказуемых и опасных охоты и собирательства к оседлости и сельскому хозяйству. В эпидемиологии этот процесс принято называть «первым эпидемиологическим переходом» (англ. First epidimiologic transition, FET).
Цель перехода в целом была достигнута. Жизнь в деревянных, а затем в каменных домах позволила более не бояться хищников. Сельское хозяйство, пусть и не со стопроцентной вероятностью, но защищало от голодной смерти. Качество и продолжительность жизни ощутимо выросли. Но на смену старым убийцам пришли новые. Дело в том, что одним из наиболее значительных следствий «перехода» стали эпидемии, которых человечество раньше не знало.
Дело в изменившемся в ходе FET образе жизни человека. До этого мы жили небольшими группами, состоявшими не более чем из 50 особей, занимавшими довольно обширные пространства. К тому же мы постоянно меняли место жительства, нигде не оставались надолго. Культура гигиены была довольно низкой — зачем следить за чистотой убежища, если ты уже съел всех мамонтов вокруг и завтра надо искать новое место?
В ходе FET люди начали надолго оставаться на одном месте, формировать более крупные группы для защиты от набегов соплеменников. Скученность и загрязнение места обитания создали оптимальные условия для развития у нас инфекций. Начались эпидемии, которые были тем свирепее, чем больше был город и чем плотнее жили в нем люди.
Новый набор «убийц» человека очень хорошо метафорически отражен в «Откровении» Иоанна Богослова (рис. 1). Силы человеческие велики, но у Бога остаются непреодолимые орудия для уничтожения рода человеческого — мор, голод и война, за которыми всегда следует смерть. И если войну можно избежать политическими мерами, к голоду можно подготовиться, то от мора можно лишь бежать со всех ног.

Рисунок 1. Васнецов В.М. «Воины Апокалипсиса», 1887.
Довольно долгое время, около 5000 лет, понадобилось человечеству, чтобы научиться справляться с инфекциями. Где-то раньше, где-то позже люди осознали важность гигиены для жизни и здоровья. Были эмпирически найдены лекарства от многих болезней. Можно сказать, весь прогресс медицины и человечества в целом происходил под постоянно довлеющим страхом новых эпидемий.
Все это не могло не отразиться на нашей культуре. Во многие мировые религии с самого начала исторического периода включались гигиенические требования. Чего стоят одни только египетские жрецы, ежедневно брившие все тело и постоянно очищавшие его от любой грязи. Слово «нечистый» во многих культурах синонимично слову «плохой», «тот, кого надо избегать». Неотделимость смерти от мора научила нас панически бояться любой нечистоты, любых признаков болезни и бежать со всех ног (рис. 2).

Рисунок 2. Обложка Le petit journal от 1 сентября 1912 года, посвященного эпидемии холеры в Индии и на Ближнем Востоке в начале 20-го века.
В настоящее время мы настолько чисты, насколько не были никогда в истории. Особенно хорошо это видно в развитых странах. Мы привыкли к тому, что во все дома подведена вода и всегда есть возможность принять ванную или душ. Мы пользуемся мылом, влажными салфетками, асептическими гелями. Мы даже моем наши дороги шампунем!
Казалось бы, ну чистые и чистые, что тут такого? При чем тут аутоиммунные заболевания? Оказывается, связь самая прямая.
Гигиеническая гипотеза
Первые наметки того, что ученые сегодня называют «гигиенической гипотезой» или «гипотезой старых друзей» появились в научной литературе еще в конце 19-го века. Этот период в истории иммунологии называют «вторым эпидемиологическим переходом» (англ. Second epidimiologic transition, SET). Он характеризуется сильным снижением заболеваемости различными инфекционными заболеваниями (бактериальными и гельминтными), а также намного более редким переходом этих заболеваний в эпидемии. Возьмем, например, чуму. Все мы из школы знаем, как сильно она влияла на судьбы Европы Средних веков. Но многие ли знают, что она отнюдь не побеждена полностью? По данным ВОЗ, в 2015 году чумой по всему миру заболели 320 человек, 77 из которых умерли. Заметьте, никакой эпидемии. Очаги оперативно локализуются, заболевшие получают адекватное лечение, почти все выздоравливают. «Черная смерть» никого больше не пугает.
Этот переход стал возможен благодаря появлению антибиотиков и других высокоэффективных противопаразитических лекарств. В развитых странах он завершился к концу 20-го века. Если в середине века в Европе каждый третий житель был поражен гельминтами [1], то в настоящий момент обнаружение носителя этих паразитов скорее редкость. Россия в этом отношении практически не отстает от развитого мира благодаря нашим сильным гигиеническим традициям. Дополнительный вклад вносит городской образ жизни, централизованное снабжение очищенной водой, контроль качества пищи и так далее.
Однако еще с 19-го века начали появляться данные, что современный городской образ жизни и общее благополучие (как правило, сопровождающееся повышенной «чистотой» жизни) приводят к определенным заболеваниям (рис. 3). Типовой аристократ викторианской эпохи обязательно имел несколько «светских» заболеваний, таких, как «сенная лихорадка» или, как мы зовем ее сейчас, аллергия на пыльцу. Более серьезным «заболеванием богатых» стал диабет первого типа [2], который был бичом «благополучного общества» до открытия целебных свойств инсулина.
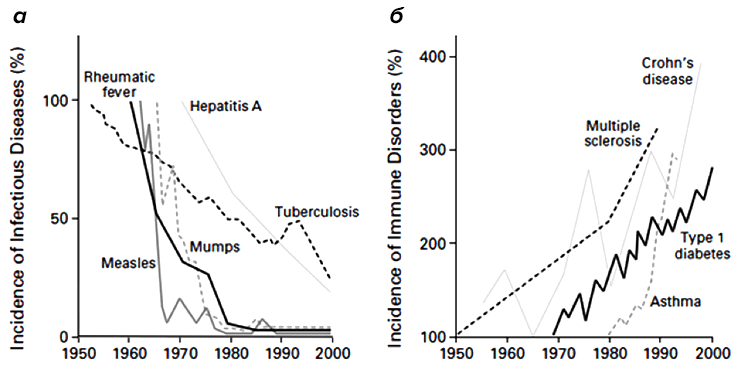
Рисунок 3. Обратное отношение между частотой инфекционных заболеваний и частотой иммунных расстройств с 1950 по 2000 годы. а — Изменение относительного количества заболевших различными инфекционными заболеваниями. б — Относительный рост заболеваемости аутоиммунными заболеваниями за тот же период.
Заболевания эти возникли в истории человечества внезапно и довольно-таки недавно — 200 лет назад они либо отсутствовали, либо были столь редки, что не оставили следа в медицинской литературе. Наличие связи между возникновением этих заболеваний и условиями жизни больных впервые свел воедино Дэвид Стракан в 1989 году в короткой заметке, где он постулировал «гигиеническую гипотезу» [3]. Он отметил, что сенной лихорадке больше подвержены те люди, у которых было меньше братьев и сестер в детстве. Стракан предположил, что устойчивость к сенной лихорадке передается с детскими инфекциями от сиблинга к сиблингу и является следствием сниженной гигиены.
Впоследствии многие исследователи показали то же самое на примере других аллергий и аутоиммунных реакций. К примеру, если детям из неблагополучных по гигиене регионов вроде Чили или Тайланда провести европейскую программу дегельминтизации, у них букетом высыпают аллергии [4].
Наверное, самым интересным примером тут является история с рассеянным склерозом [5–8]. Ученые решили посмотреть, что происходит, когда больной этим страшным заболеванием заражается гельминтами, и начали искать инфицированных червями пациентов с РС. Результаты были ошеломляющи. У пациентов, которые заражались определенными гельминтами (например, власоглавом Trichuris trichiura) течение заболевания практически останавливалось [6]. Во время инфекции у них на 95% снижалось количество новых бляшек в мозге (рис. 4). Результат, недостижимый ни одним современным методом терапии! Если же по каким-то причинам гельминтов требовалось удалить (например, развивалось острое воспалительное поражение кишечника), болезнь возобновлялась с той стадии, на которой остановилась при инфекции.
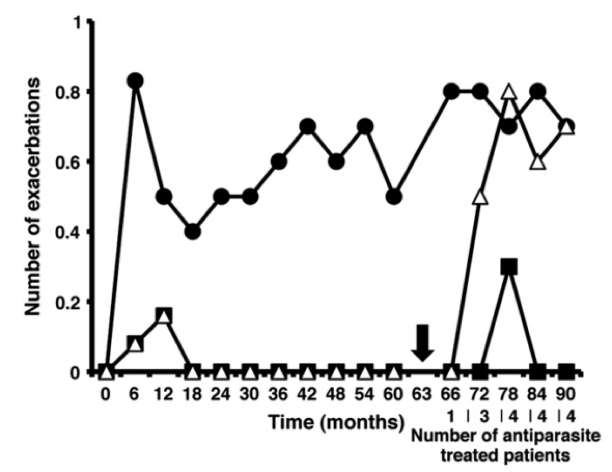
Рисунок 4. График появления новых бляшек в мозге (когортное исследование 2011 года). Круги — неинфицированные гельминтами пациенты, квадраты — инфицированные, треугольники — инфицированные, но вылеченные от гельминта (момент излечения показан черной стрелкой).
Можно сказать, власоглав поддерживал здоровье этих людей, позволяя им вести нормальный образ жизни, пока они позволяют ему жить внутри себя. Так что же происходит? Как черви справляются с задачей, с которой не способна справиться современная медицина? Для ответа на этот вопрос нам придется разобраться, как же работает наш иммунитет.
Иммунный ответ
Иммунная система призвана защищать организм от внутренних и внешних врагов. Внешними врагами являются вирусы, бактерии, простейшие и черви, которые постоянно попадают в наш организм и уничтожаются на дальних рубежах. Внутренними врагами являются раковые клетки, а также клетки, зараженные вирусами или внутриклеточными бактериями.
Ключевые для иммунитета понятия — «антиген» и «воспаление». Антиген — это какая-либо молекула, которую способна узнать и атаковать иммунная система. Практически что угодно может быть антигеном. Воспаление же — это реакция ткани на повреждение или опасность такого повреждения. Молекулы, запускающие воспаление, называются провоспалительными, а блокирующие его — противовоспалительными.
Когда паразит попадает в организм, первым делом его встречает врожденный иммунитет, клетки которого (макрофаги) есть во всех тканях. Антигеном в данном случае служат нехарактерные для нашего организма молекулы — клеточная стенка бактерий, двухцепочечная РНК некоторых вирусов, свободно плавающая в межклеточном пространстве наша ДНК и так далее. При обнаружении пришельцев, клетки врожденного иммунитета пытаются их уничтожить, параллельно выделяя провоспалительные молекулы (рис. 5). Воспаленная ткань блокирует выход паразита из места проникновения в остальной организм и привлекает новые клетки иммунитета к месту повреждения.
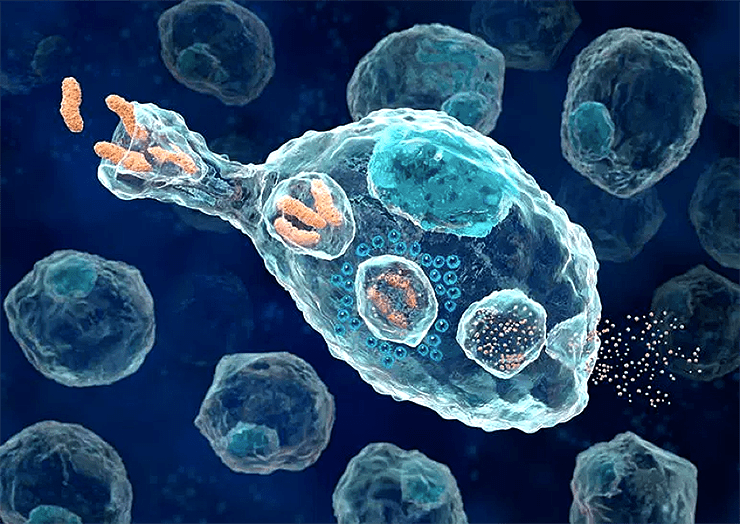
Рисунок 5. Фагоцит пожирает бактерии.
Если врожденному иммунитету не удается уничтожить захватчиков, в дело вступает адаптивный иммунитет. Происходит это отнюдь не сразу: активации адаптивного ответа предшествуют 3–4 дня подготовки в лимфоузлах (при этом лимфоузлы увеличиваются в размерах, что является признаком инфекционного заболевания). Начинается все с того, что некоторые из клеток врожденного иммунитета прибывают в лимфоузел, неся на себе антигены из места поражения. Антигенами в данном случае выступают короткие (от 8 до 20 аминокислот) пептиды из белков инфекционного агента и окружающих тканей. По сути, макрофаг (или специализированный активатор адаптивного иммунитета — дендритная клетка) просто захватывает из места воспаления образцы растворенных белков, ошмётков паразита и погибших клеток и приносит в лимфоузел.
В лимфоузле его встречают наивные (неактивированные) клетки адаптивного иммунитета — Т-лимфоциты. Каждый лимфоцит, выйдя из места своего формирования, несет на себе уникальный рецептор, который формируется путем направленного внесения мутаций в геном. Заранее неизвестно, может ли этот рецептор распознать какой-либо антиген, но его вариантов так много (по некоторым оценкам, у нас может быть до 10 48 разных типов этого рецептора, но большая часть их будет нефункциональна), что в течение нескольких часов в лимфоузле обнаруживается как минимум несколько клеток, способных распознать антигены паразита. Затем эти клетки делятся, активируются и отправляются в поврежденную ткань, где отыскивают свои антигены и уничтожают как самих захватчиков, так и зараженные клетки, если мы говорим о вирусе или внутриклеточной бактерии (рис. 6).
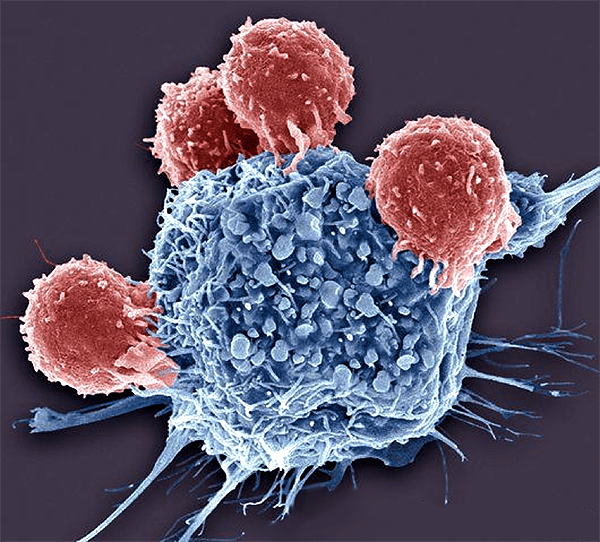
Рисунок 6. Т-лимфоциты (красные) убивают раковую клетку (синяя) своего же организма. После получения сигнала раковая клетка начинает распадаться на небольшие пузырьки, которые затем съедят клетки врожденного иммунитета.
Иммунологическая толерантность
Иммунная система — единственная из систем организма, в чью задачу входит уничтожение других живых существ — отдельных клеток или многоклеточных организмов. Причем наши собственные клетки тоже часто должны уничтожаться, если они заражены вирусом, бактерией или превратились в раковые. При этом необходимо избегать иммунного ответа на нормальные клетки. Если такой ответ развивается — возникает аутоиммунное заболевание.
Чтобы этого избежать, в нашем организме есть система создания иммунологической толерантности — защиты «своего» от иммунитета. Центральная толерантность заключается в уничтожении в процессе развития тех Т-лимфоцитов, которые мутировали свои рецепторы так, что они могут узнать и атаковать свои антигены. Часть таких лимфоцитов из убийц превращается в защитников (так называемые регуляторные Т-лимфоциты) — они узнают заведомо «свой» антиген и подавляют любой иммунный ответ против него.
Периферическая толерантность возникает, когда Т-лимфоцит распознает антиген в лимфоузлах, но никакого воспаления в месте, откуда этот антиген попал в лимфоузел, нет. Напротив, высока концентрация противовоспалительных молекул. Такой лимфоцит опять-таки или уничтожается, или превращается в регуляторный.
Паразиты и симбионты
Миллиарды лет эволюции крупные многоклеточные организмы были домом и едой для более мелких одноклеточных и многоклеточных. Человек тут не исключение — ведь мы являемся как хорошим источником пищи, так и отличным защитником для всего, что сумеет поселиться внутри нас или на нас.
Эволюция поделила этих сожителей на 2 большие группы — паразиты и симбионты. Паразиты делают ставку на быстрое размножение. У них есть возможность подавления врожденного иммунитета, а пока адаптивный активируется, они уже успевают размножиться за счет наших ресурсов и передать инфекцию дальше. Так действует, например, вирус гриппа или бактериальная пневмония.
Симбионты же научились подавлять как врожденный, так и адаптивный иммунитеты. Для этого им пришлось умерить свои аппетиты — если клетки организма постоянно повреждаются, то никакие уловки не смогут предотвратить активацию иммунитета. Потому они поселились на поверхностях нашего тела, прежде всего на поверхности ЖКТ, где они получают лишь часть нашей пищи, но не покушаются на сам организм.
Помимо этого, они научились подавлять воспаление, выделяя вещества, которые похожи на наши противовоспалительные молекулы. Макрофаги врожденного иммунитета, столкнувшись с такими бактериями, могут почувствовать антигены клеточной стенки, но не активируются, так как подавлены противовоспалительным фоном вокруг.
Третьим механизмом защиты стала антигенная мимикрия. Для адаптивного иммунитета основным антигеном являются пептиды из белков. И многие наши симбионты в ходе эволюции поменяли свой белковый состав так, чтобы в нем был максимум пептидов, похожих на наши. Таким образом они встают под защиту регуляторных лимфоцитов. Этот механизм характерен для всех видов наших сожителей — бактерий, червей (рис. 7), вирусов и так далее.
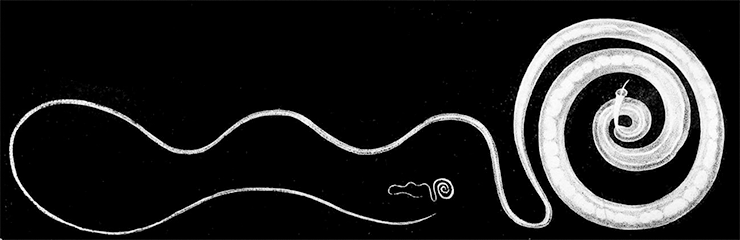
Рисунок 7. Власоглав — один из гельминтов, активно изучаемых в рамках гигиенической гипотезы.
В течение миллионов лет каждая особь нашего вида, рождаясь, сразу же вступала в контакт с симбионтами, населявшими кожу, слизистые и кишечники своих собратьев. Со временем организм научился извлекать выгоду из такого постоянного неустранимого сосуществования. В частности, способность бактерий и червей создавать сильный противовоспалительный фон в месте своего обитания стала за это время ключевым фактором создания периферической толерантности. Она распространилась как на антигены самих сожителей, так и на сопутствующие им — антигены пищи (в кишечнике), пыли и пыльцы (в легких) и собственного организма (те самые антигены, которые сожители развили в ходе антигенной мимикрии).
И снова гигиеническая гипотеза
Внимательный читатель уже смог уловить, где тут связь. Особенность второго эпидемиологического перехода состоит в том, что мы с вами чисты, как никогда прежде, лишены практически всех червей и многих патогенов. Ребенок видит свою первую грязь или лужу отнюдь не в первые дни жизни, как раньше. Антибиотики и правила гигиены, центральное водоснабжение и мытье асфальта шампунем несомненно сделали нашу жизнь лучше. Но незаметно для себя вместе с «грязью» мы также начали устранять из своей жизни и часть тех самых симбионтов, червей и бактерий (а по мнению некоторых ученых — и некоторых вирусов), которые помогали нам создавать толерантность к собственному организму и аллергенам окружающей среды.
Уже сейчас мы имеем доказательства того, что заражение некоторыми видами симбиотической флоры ведет к снижению частоты многих аутоиммунных заболеваний [9], [10]. Это как раз та самая флора, которая широко представлена в странах третьего мира и почти не представлена в развитых странах. Пример с «остановкой» рассеянного склероза лишь самый яркий, но таких примеров намного больше. Показано, что у пациентов, зараженных таким ныне изгнанным «паразитом», повышается количество регуляторных Т-клеток, возрастает концентрация противовоспалительных молекул [9]. Изгнание симбионта возвращает все вспять. У некоторых симбионтов выявлена сильная связь с диабетом, у других — с рассеянным склерозом и так далее.
Особенно сильно на риск развития таких заболеваний влияет первый год жизни. Если в этот период ребенок оказывается в деревне, проводит некоторое время в больших группах сверстников (в больнице или в детском саду) и вообще чаще встречается с инфекциями — риск развития аутоиммунных заболеваний серьезно снижается [11], [12].
Разумеется, не только микробиом (совокупность всех симбиотических микроорганизмов конкретного человека) [13] и гельминты влияют на риск развития аутоиммунных и аллергических реакций. Есть и генетическая предрасположенность, и условия, в которых человек впервые встречается с тем или иным внешним антигеном. Есть некоторые микроорганизмы, которые не защищают, а, напротив, провоцируют аутоиммунные заболевания. Например, стрептококк способен вызывать ревматизм, а некоторые стафилококки производят суперантиген, который неспецифически запускает все клоны Т-лимфоцитов с любым рецептором — это тоже может привести к аутоиммунным заболеваниям.
Но исключения лишь подтверждают правило. Способность симбионтов и паразитов влиять, позитивно или негативно, на развитие аутоиммунных заболеваний — уже доказанный факт. Что делать с этой информацией, врачи и ученые пока не знают. Мы пробовали получать гомогенаты червей и использовать их в качестве лекарства [14]. Это не сработало. Иммунологи выясняют механизмы, с помощью которых черви достигают того, что недоступно всем врачам мира, а врачи и фармацевтические компании разрабатывают инновационные методы терапии. По состоянию на 2015 год во всем мире проводилось свыше 20 клинических испытаний «гельминтной терапии», в ходе которой пациенты принимают дозированную, полученную в стерильных условиях лучших фармпроизводств суспензию живых яиц глистов (табл.) [15]. Несмотря на неплохие результаты [16], проблемы все же остаются [4]. Например, часто гельминтов приходится удалять из-за развивающегося воспалительного заболевания кишечника. Но лучшего варианта у нас пока нет.
| Заболевание | Количество исследований | Общее количество пациентов | Результаты |
|---|---|---|---|
| Болезнь Крона | 6 | 543 | Показана безопасность и статистически значимые улучшения у большинства пациентов. |
| Неспецифический язвенный колит | 3 | 192 | Показана безопасность и статистически значимые улучшения у большинства пациентов. |
| Рассеянный склероз | 6 | 156 | Показана безопасность, статистически значимые улучшения у части пациентов. |
| Непереносимость глютена (целиакия) | 2 | 35 | Исследования только начались |
| Расстройства аутического спектра | 3 | 90 | Пилотное исследование показало эффективность, подтверждающие только начаты |
| Псориаз | 3 | 55 | Исследования только начались |
| Аллергия на арахис | 1 | 18 | Исследования только начались |
| Бронхиальная астма | 1 | 32 | Зафиксированы статистически недостоверные улучшения |
| Аллергический риноконъюнктивит | 2 | 130 | Эффективность не показана |
| Ревматоидный артрит | 1 | 50 | Исследование только началось |
О ревматоидном артрите рассказано в статье «Ревматоидный артрит: изменить состав суставов» [17], а псориазу на «Биомолекуле» посвящен целый спецпроект — «Псориаз». — Ред.
Что же делать нам, простым смертным, пока ученые по кусочкам разбирают эту тайну и ищут решение? Начать стóит со снижения маниакального стремления к чистоте во всем. Я не предлагаю не мыть руки перед едой. Но довольно часто в последнее время мы перегибаем палку. Антибиотики при каждом чихе, асептические спиртосодержащие гели каждые 10 минут, антибактериальное мыло вместо обычного в ванной. Все эти меры способны спасти вас от эпидемии. Но каждодневное их применение, особенно детьми, способно нанести куда больший урон, чем грипп или пищевое отравление.
Источник https://ru.abcdef.wiki/wiki/Plant_tolerance_to_herbivory
Источник https://biomolecula.ru/articles/starye-druzia-kliuch-k-autoimmunnym-zabolevaniiam
Источник
Источник