Почему болеют растения и как они борются с инфекцией на примере фитопатогенных бактерий
Содержание
Почему болеют растения и как они борются с инфекцией на примере фитопатогенных бактерий
 Обзор
Обзор
Сосудистый бактериоз капусты (справа).
Автор
Редакторы
- Биология
- Гормоны растений
- Микробиология
Статья на конкурс «Био/Мол/Текст»: Растения болеют. Наверное, этот тезис у читателя не вызовет какого-то особенного удивления. Данный факт кажется весьма очевидным, учитывая, что многие обыватели любят выращивать растения у себя дома или в саду и с большой вероятностью сталкивались с подобной проблемой. Однако, что может быть не совсем очевидным, так это то, что растения обладают своеобразной иммунной системой, защищающей их организм от внедрения различных патогенов — грибов, бактерий, вирусов, круглых червей. Да-да, все прямо как у людей, или нет? Давайте разбираться.

Конкурс «Био/Мол/Текст»-2020/2021
Эта работа опубликована в номинации «Свободная тема» конкурса «Био/Мол/Текст»-2020/2021.
Генеральный партнер конкурса — ежегодная биотехнологическая конференция BiotechClub, организованная международной инновационной биотехнологической компанией BIOCAD.

Спонсор конкурса — компания SkyGen: передовой дистрибьютор продукции для life science на российском рынке.
Спонсор конкурса — компания «Диаэм»: крупнейший поставщик оборудования, реагентов и расходных материалов для биологических исследований и производств.
Инфекционные болезни растений и их возбудители
Болезни растений принято делить на неинфекционные и инфекционные. Причинами первых является недостаток или избыток того или иного фактора неживой природы — воды, света, минеральных веществ и т.д. Вторые вызываются различными микроорганизмами — бактериями, вирусами, грибами, круглыми червями-нематодами [1].
С болезнями растений человечество сталкивалось с незапамятных времен, и порой подобные «столкновения» заканчивались для нас не самым лучшим образом.
Например, эпидемии «Антониевого огня» в Средние века были вызваны массовыми отравлениями людей токсинами грибка-спорыньи (Claviceps purpurea), паразитирующего на пшенице и ржи (рис. 1) [2].

Рисунок 1. Антониев огонь или эрготизм — заболевание, вызываемое отравлением алкалоидами гриба спорыньи (Claviceps purpurea).
(а) — склероции спорыньи на колосе злакового растения, содержащие большое количество алкалоидов.
(б) — больной эрготизмом, изображенный на Изенгеймском алтаре художником Маттиасом Грюневальдом (1506–1515 гг.). Болезнь была крайне распространена в Средневековье в Европе и в России, однако встречалась вплоть до XX века (известна вспышка в 1951 году во Франции) [2]. Масштабное распространение эрготизма объяснялось низкими требованиями к качеству зерна, которое в сырые и холодные годы активно поражалось спорыньей. Алкалоиды спорыньи (в особенности эрготин) при попадании в организм вызывают длительные спазмы гладкой мускулатуры, судороги, расстройства нервной системы, а при больших дозах — смерть. У эрготизма существует две формы: гангренозная (известная как «Антониев огонь») и конвульсивная ( «ведьмина корча»).
коллаж автора статьи по материалам «Википедии» и сайта aokin
Голод в Ирландии в 1845–1849 годы, в свою очередь, стал следствием масштабной гибели посевов картофеля по причине заражения оомицетом фитофторой (Phytophthora infestans) [3].
Грибковые заболевания растений преследуют человечество и по сей день, являясь, наверное, одной из самых распространенных причин гибели посевов в сельском хозяйстве. Помимо спорыньи и фитофторы, возбудителями заболеваний растений являются представители родов Fusarium, Alternaria, Microdochium, Typhula и других [4].
В зависимости от стратегии, к которой прибегают грибы для «захвата» растения- хозяина, их, как и все фитопатогены в целом, принято разделять на три группы — некротрофы, биотрофы и гемибиотрофы. Некротрофы убивают клетки растительных тканей, после чего заселяют пораженный участок и поглощают из него питательные вещества, биотрофы же извлекают необходимые вещества непосредственно из живых клеток. Гемибиотрофы — промежуточный вариант: сначала они питаются биотрофно, а затем, после гибели зараженных тканей, продолжают жить на них как некротрофы [1]. Внешние проявления фунгиозов крайне разнообразны: от различного рода бурых пятен, ржавчины или белоснежного налета на листьях до гниения корней и масштабных некрозов отдельных органов [5].
Не менее часто растения становятся жертвами бактериозов — заболеваний, вызываемых патогенными бактериями. Например, Pseudomonas syringae — возбудитель базального бактериоза зерновых — один из самых распространенных фитопатогенов в Европейской части России: он способен поражать до 70% урожая некоторых сортов пшеницы и ячменя.
Стратегии колонизации бактериями организма хозяина могут быть достаточно разнообразными: от грубого разрушения клеток литическими ферментами до искусного внедрения в геном растения части своей плазмидной ДНК. Внешние проявления бактериозов самые различные — мокрые гнили, бурые пятнистости, увядание, образование опухолей [5].
Растения, подобно животным и бактериям, могут также поражаться и вирусными частицами. Интересно, что первым открытым вирусом стал вирус табачной мозаики (Tobacco mosaic virus) — фитопатоген, паразитирующий на представителях семейства пасленовых (табак, картофель и т.д.). Вирусы проникают в клетки растения через уже имеющиеся повреждения, после чего происходит высвобождение и репликация вирусной нуклеиновой кислоты и синтез вирусных белков на рибосомах организма-хозяина. Внутри растения вирусы распространяются либо по сосудистой системе, либо через плазмодесмы (специальные соединительные мостики между клетками растений) [6]. Внешними характерными признаками вирусных заболеваний у растений могут быть мозаичная окраска, желтушность или скручивание листьев, их неравномерный рост, иногда — усиление ветвления и укорочение междоузлий (рис. 2) [4].

Рисунок 2. Листья табака, пораженного вирусом табачной мозаики.
Кроме грибов, бактерий и вирусов, на растения охотятся еще и круглые черви. Наиболее вредоносными нематодами в мире считаются виды родов Meloidodyne, Heterodera, Uobodera, Ditylenchus,Tylenchulus [7].
Практически все известные нам высшие растения могут быть потенциальными хозяевами одного или более видов паразитических червей. В ротовой полости фитопатогенных нематод расположен стилет — специальный колюще-сосущий орган, которым нематода прокалывает ткани растений и высасывает из них питательные вещества, вызывая образование галл или язв на корнях (галловые нематоды), деформацию и некроз листьев (листовые нематоды), общее угнетение роста и развития организма хозяина (рис. 3). Помимо этого, нематоды зачастую переносят на себе болезнетворные грибы, вирусы и бактерии, способные вызывать сопутствующую инфекцию [8].

Рисунок 3. Внешние проявления поражения растений нематодами.
(a) — нематода (Globodera rostochiensis) как пример фитопатогенного круглого червя;
(б) — внешние проявления поражения растений нематодами: защитные цисты золотистой нематоды на корнях картофеля;
(в) — внешние проявления поражения растений нематодами: галлы на корнях пораженного галловыми нематодами паслена;
(г) — засохшие кончики листьев и порча семян у риса;
(д) — бурые пятна на листьях папоротника.
Каждому растению — свой патоген
Не стоит думать, что любой фитопатоген способен инфицировать любое растение. Для образования так называемой патологической системы (системы взаимодействия паразит-хозяин) оба ее участника должны обладать генетической и физиологической совместимостью. Это значит, что для успешной интеграции растение и патоген обязаны обладать соответствующими наборами генов, продукты которых обеспечат их успешное узнавание и взаимодействие; также оба они должны находиться в определенном физиологическом состоянии, которое зависит от стадии развития организма и внешних условий [1], [9].
Бактериозы растений
Поскольку мы будем рассматривать тему фитоиммунитета на примере бактериальных инфекций, о последних стоило бы рассказать подробнее.
Бактериозы растений считаются животрепещущей проблемой мирового сельского хозяйства XXI века. Долгое время опасность бактерий недооценивали, отдавая пальму первенства в этом вопросе грибам. Тем не менее, как было сказано ранее, фитопатогенные бактерии способны вызывать развитие серьезных патологий у растений, таких как бактериальный рак, мягкие гнили (рис. 4), некрозы, ожоги и увядания отдельных органов, потенциально приводящие к смерти целого организма. Наиболее опасными фитопатогенными бактериями являются представители родов Pectobacterium, Xanthomonas, Pseudomonas, Dickeya, Agrobacterium и другие.
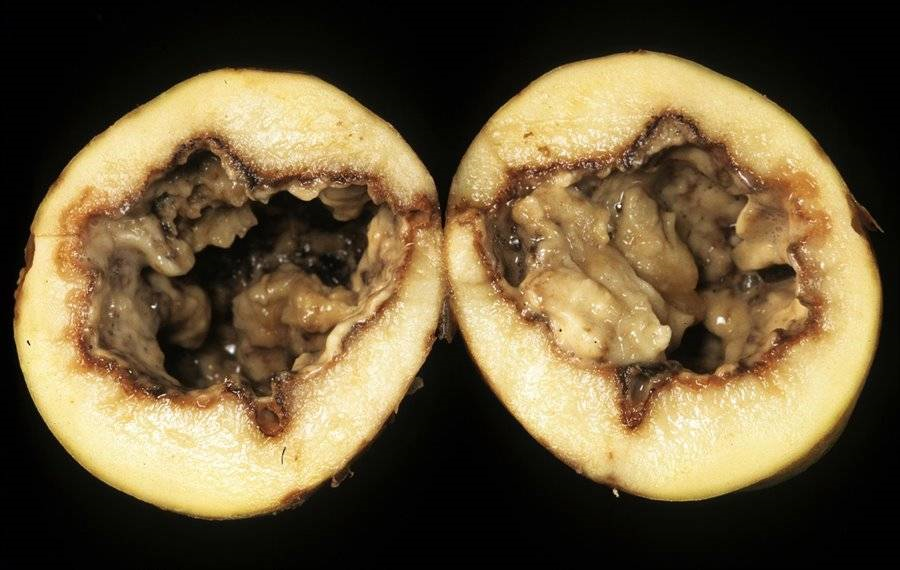
Рисунок 4. Мягкая гниль картофеля, вызванная фитопатогенной бактерией Pectobacterium carotovorum.
У бактерий существует ряд инструментов, с помощью которых они колонизируют организм растения-хозяина. Первое оружие — это наличие у фитопатогенных бактерий огромного разнообразия выделяемых наружу литических ферментов, разрушающих растительную клеточную стенку. Второе — фитотоксины, особые низкомолекулярные вещества, направленные на убийство растительных клеток или же подавление иммунного ответа хозяина. Третье — сидерофоры, тоже низкомолекулярные соединения, необходимые бактериям для транспорта железа из внешней среды внутрь клетки. Бактерии используют железо для синтеза веществ, позволяющих им успешно развиваться в организме растения. Четвертое — жгутики и пили, обеспечивающие подвижность микроорганизмов. Пятое — экзополисахариды, которые выполняют множество функций: структурную (объединяют бактерии в единые структуры — биопленки); адгезивную (позволяют бактериям прикрепляться к субстрату, что особенно важно при колонизации фитопатогенами сосудов ксилемы, где у растений идет транспорт воды); защитную (предохраняют клетки бактерий от высыхания, от активных форм кислорода, а также могут работать наподобие стелс-технологии, скрывая от защитных систем организма растения патогенные детерминанты, например, флагеллин — белок жгутиков). У некоторых патогенных микроорганизмов, например, у агробактерий, имеются специальные системы, позволяющие им встраивать часть своей ДНК в геном растения, тем самым заставляя последнее синтезировать необходимые фитопатогену вещества [9].
Как видно, военный арсенал у фитопатогенных организмов действительно крайне разнообразен. А что же растения? Поверьте, им тоже есть, чем ответить.
Как растение борется с «заразой» на примере бактериальных инфекций
В отличие от животных, у растений нет специальных иммунных клеток (лейкоцитов) для того, чтобы противостоять фитопатогенам. Почти все клетки растения способны запускать иммунный ответ и защищать себя с помощью так называемых факторов устойчивости. Что же это такое?
Факторы устойчивости — это особые структуры, метаболиты, сигнальные системы, существующие в растении и участвующие в защите последнего от фитопатогенов.
Здесь стоит отметить, что культурные растения в целом гораздо более подвержены заражению различными фитопатогенами по сравнению с своими «дикими родственниками» в том числе именно потому, что селекция новых сортов часто идет вразрез с желанием растения синтезировать разнообразные защитные метаболиты.
Факторы устойчивости принято делить на конститутивные и индуцируемые. Под конститутивными факторами подразумеваются особые барьерные структуры клетки: целлюлозная клеточная стенка, наружная кутикула, покрывающая листья и стебли многих растений, а также постоянно синтезируемые в растении особые вещества, губительно действующие на незваных гостей. Индуцированный же иммунный ответ запускается растением исключительно в присутствии того или иного патогена. Организм понимает, что в него попал кто-то «чужой» благодаря специальным рецепторам, расположенным внутри или снаружи клетки. Они распознают особые «сигналы опасности» и активируют защитные системы. Подобными сигналами (элиситорами) могут быть консервативные молекулы, являющиеся компонентами клеточных структур фитопатогенов, разнообразные вещества (факторы вирулентности), секретируемые паразитами наружу (в апопласт), или «обломки» собственных полимеров растения, которые разрушаются в процессе колонизации патогенами организма хозяина. Элиситоры бывают специфичными, то есть характерными чуть ли не исключительно для конкретного штамма, и неспецифичными, свойственными для многих фитопатогенов.
Индуцированная устойчивость бывает локальной и системной. Локальная устойчивость — это по сути индуцированная устойчивость, которая развилась строго в том месте, где произошло попадание патогена в организм растения. С биохимической точки зрения она может проявляться в усилении синтеза активных форм кислорода, защитных белков или фитоалексинов. Не стоит путать с реакцией гиперчувствительности, речь о которой пойдет позже; она является лишь частным проявлением локальной устойчивости! Системная устойчивость развивается в результате физиологических изменений, происходящих в неинфицированных клетках во всем организме растения в результате локальных изменений, вызванных инвазией патогена, и тесно связана с синтезом специальных антимикробных белков [9].
Раунд 1. Растительная клеточная стенка — первый барьер на пути непрошенных гостей
Растительная клеточная стенка (РКС) — достаточно сложно устроенная структура. И недаром! Именно она «стоит на страже» клеточного метаболизма и является первым форпостом на пути непрошенных гостей. РКС состоит из трех основных компонентов — микрофибрилл целлюлозы, связующих гликанов и пектиновых веществ (рис. 5) [10].
На первый взгляд, растительная клеточная стенка воистину неприступна, и в какой-то степени это действительно так. Далеко не все фитопатогены, потенциально способные заразить растение, прорываются через нее. Вирусы, например, в принципе не способны проникать сквозь РКС; они попадают внутрь клетки только в том случае, если она повреждена по каким-либо другим причинам. Бактерии сами по себе также не могут ее преодолеть, поскольку поры между фибриллами целлюлозы крайне малы.
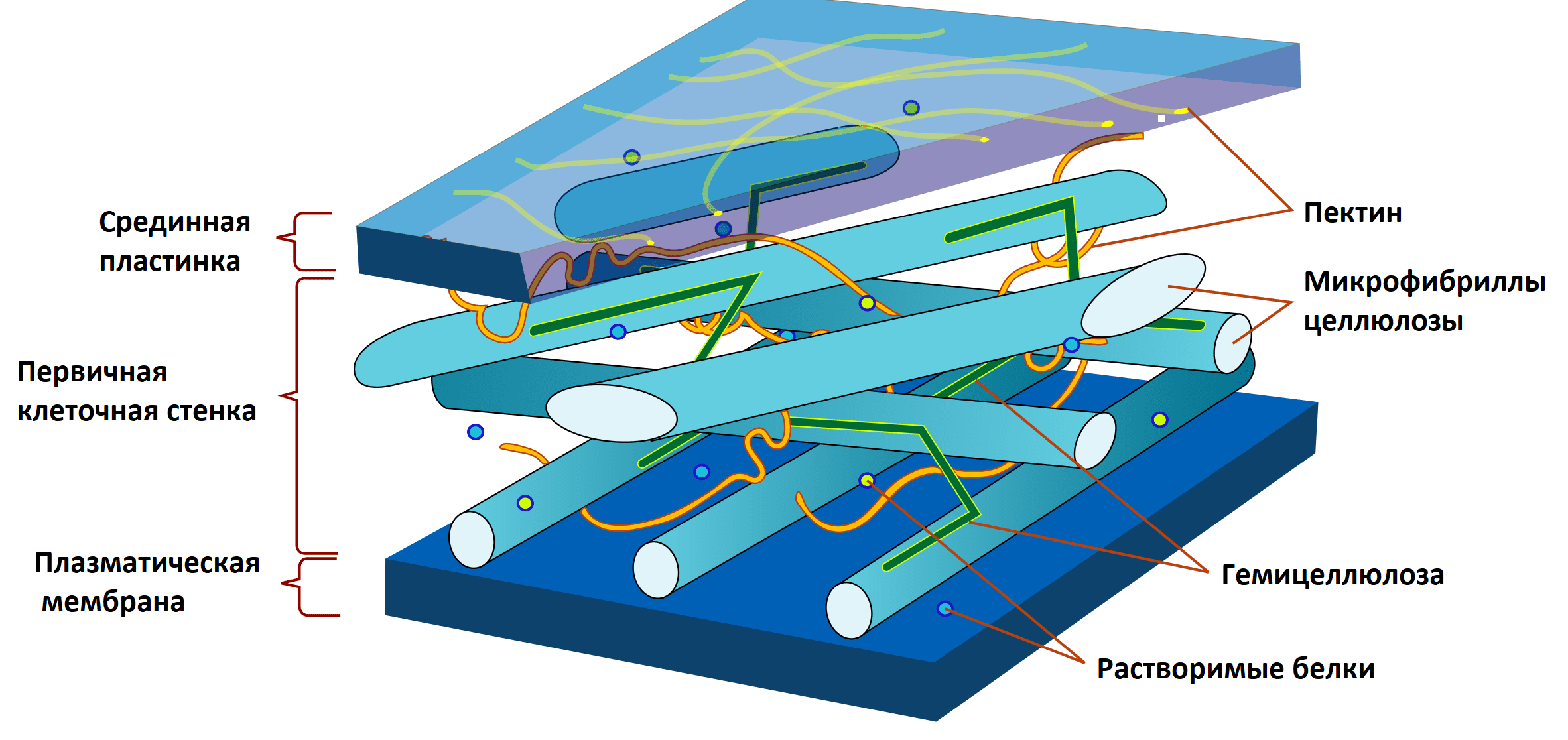
Рисунок 5. Строение растительной клеточной стенки. Целлюлоза представляет собой углеводный полимер, мономером которого является глюкоза, чьи цепочки собраны в прочные фибриллы, расположенные в виде перекрывающихся слоев. Они соединены между собой специальными углеводными «склейками» — связующими гликанами, которые называют гемицеллюлозами. Более того, вся описанная выше конструкция, как в «желе», погружена в матрикс из пектиновых веществ. Стоит отметить, что, помимо основных компонентов, РКС насыщена ионами кальция, а также содержит огромное количество разнообразных белков.
Если нельзя протиснуться сквозь барьер, значит, его нужно разрушить! Именно это зачастую и случается с РКС, когда в дело вступают специализированные фитопатогены, секретирующие огромное количество литических ферментов, позволяющих уничтожить, казалось бы, неприступную «крепость». Но нельзя сказать, что растение на этом этапе сдается. РКС может быть заведомо укреплена специальными веществами, такими как лигнин, суберин и кутин, образующими гидрофобный слой, доставляющий фитопатогенам дополнительные проблемы. К тому же лигнин и суберин можно отнести еще и к факторам индуцированной устойчивости, потому что синтез этих веществ активируется непосредственно в ответ на присутствие микроба [10], [11].
Раунд 2. Вторичные метаболиты
Что ж, клеточная стенка разрушена, патоген ворвался внутрь клетки, и именно здесь начинается следующий раунд — встреча с вторичными метаболитами.
Вторичные метаболиты — это «дополнительные» химические соединения, не участвующие в основном клеточном обмене. Они могут присутствовать не во всех клетках организма, однако имеют при этом важное функциональное значение на уровне целого растения. Вторичные метаболиты — незаменимые помощники растений. Они способны выполнять самые разнообразные функции: защищать от фитопатогенов, участвовать в размножении растений и их взаимодействии с другими организмами и т. д.
Не каждое вещество, синтезируемое растением, можно назвать вторичным метаболитом. Для них существует несколько критериев:
- выраженная биологическая активность;
- небольшая молекулярная масса
- ограниченное число предшественников их биосинтеза.
На самом деле, вторичные метаболиты способны синтезировать и животные, но растениям в этом плане нет равных! Они являются бесспорными чемпионами по количеству и разнообразию таких веществ. К слову, именно вторичными метаболитами определяются лекарственные свойства растений [9].
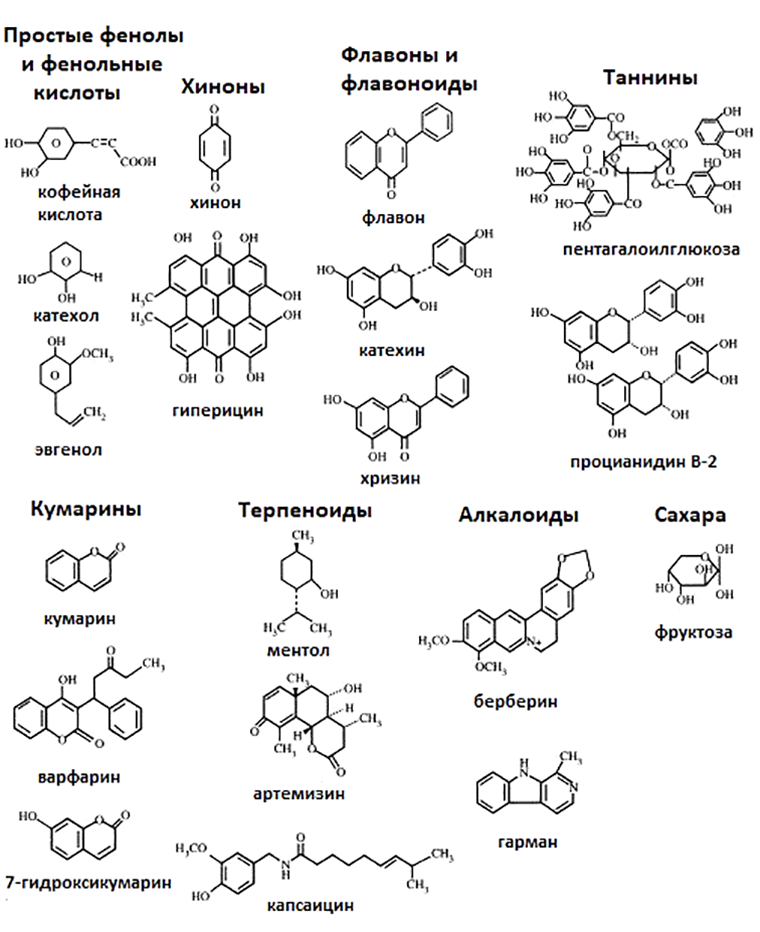
Рисунок 6. Химические структуры разнообразных вторичных метаболитов растений, обладающих антимикробными свойствами.
Наиболее изученными из данных веществ в плане фитоиммунитета являются фенольные соединения, алкалоиды и изопреноиды (терпеноиды). Все они обладают бактерицидным действием и воздействуют на широкий спектр патогенов (рис. 6). Основными мишенями вторичных метаболитов в бактериальной клетке, как правило, являются мембраны, различные белки и нуклеиновые кислоты [12].
Например, фенольные соединения (вещества ароматической природы) способны встраиваться в мембраны бактерий, приводя к их разрыву, инактивировать белки, влиять на работу их электрон-транспортных цепей, нарушая тем самым синтез АТФ, служить в качестве акцепторов электронов и окислять разные группы соединений, а также сшивать полимеры РКС, создавая тем самым дополнительные преграды на пути патогена [13].
Среди фенолов флавоноиды (окисленные формы фенольных соединений) наиболее знакомы нам, простым обывателям. Мы с удовольствием употребляем их вместе с едой и напитками растительного происхождения: с лимонами, луком, зеленым чаем, красным вином и даже черным шоколадом. Считается, что флавоноиды оказывают положительный эффект на наше здоровье. Например, известно, что флавоноиды цитрусовых снижают риск развития сердечно-сосудистых заболеваний и диабета, поскольку обладают антиоксидантными свойствами, подавляют процесс воспаления, улучшают толерантность к глюкозе и чувствительность к инсулину, модулируют метаболизм липидов [14].
Алкалоиды же — представители другой большой группы азотсодержащих вторичных метаболитов растений; они оказывают губительное влияние на синтез нуклеиновых кислот и репарацию ДНК бактерий, а также выступают в качестве ингибиторов синтеза аминокислот [15]. К алкалоидам относится такое хорошо известное вещество, как никотин: он необходим растениям для защиты от насекомых, а человеком успешно применяется как инсектицид.
Принцип действия терпеноидов на данный момент не выяснен до конца, однако считается, что он связан с нарушением структуры и функций бактериальных мембран, поскольку эти соединения являются липофильными. Известно, что они активны против бактерий, вирусов, грибов и протозоев. Кстати, именно наличием терпеноидов объясняется антимикробный эффект эфирных масел [12].
Также стоит отметить, что, в зависимости от времени синтеза, вышеупомянутые антимикробные вторичные метаболиты принято делить на три группы: фитонциды — образуются в клетках растений постоянно (конститутивный иммунитет); фитоалексины — синтезируются только в присутствии фитопатогена и являются частью индуцированного ответа; полуиндуцибельные — до внедрения патогена хранятся в клетке в неактивной форме [9].
Таким образом, огромное разнообразие вторичных метаболитов и многообразные механизмы их действия позволяют растениям эффективно противостоять непрошенным гостям. Эти метаболиты способны выводить из строя жизненно важные системы микроорганизмов, а также способствовать укреплению барьеров растительной клетки.
Раунд 3. Индуцированный иммунный ответ
Хотя факторы конститутивной устойчивости и способны препятствовать проникновению в растение большинства потенциальных патогенов, ряд микроорганизмов может преодолевать подобные преграды. Здесь вступают в бой особые сигнальные системы растения, способные распознать чужеродные вещества (элиситоры) и активировать адекватный ответ на них.
Локальная устойчивость
Мы уже знаем, что, в зависимости от локализации и механизма иммунного ответа, различают локальную и системную устойчивость. Локальную устойчивость можно разделить еще на два типа:
- количественная устойчивость (патоген-ассоциированный молекулярный паттерн (ПАМП)-индуцированная или горизонтальная устойчивость);
- качественная устойчивость (эффектор-индуцированная или вертикальная устойчивость).
Количественная устойчивость считается эволюционно более древним типом индуцированной устойчивости, характерным для всех семенных растений. Она называется количественной, поскольку в процессе ее «работы» болезнь у растения может проявляться в той или иной степени или не проявляться совсем — тогда растение будет абсолютно устойчивым к фитопатогену. Количественная устойчивость абсолютно не специфична и участвует в защите организма от широкого спектра фитопатогенов. Вещества, способные вызывать количественную устойчивость, делятся на две основные группы:
- Метаболиты патогенных микроорганизмов, без которых существование последних было бы невозможно (например, хитин у грибов, входящий в состав их клеточной стенки, или флагеллин бактерий — основной компонент их жгутиков) — то есть в процессе эволюции патогены не могут от них избавиться.
- Продукты распада собственных растительных полимеров, которые сигнализируют растению об опасности. Ими могут быть, например, олигосахариды, образующиеся в процессе разрушения РКС.
Рецепторы количественной устойчивости, распознающие внешние и внутренние «сигналы опасности», расположены на мембране клетки. Они, как правило, либо сами обладают киназной активностью, либо связаны с внутриклеточными киназами — ферментами, присоединяющими остатки фосфора к другим белкам.
Механизм работы количественной устойчивости на примере элиситора флагеллина
Давайте разберемся, как работает локальная количественная устойчивость, на примере флагеллин-чувствительного рецептора (FLS2), впервые описанного у резуховидки Таля и имеющегося у большинства исследованных видов растений. Рецептор распознает консервативный фрагмент флагеллина, после чего передает сигнал внутрь клетки. На присутствие бактериального белка растительная клетка реагирует несколькими способами (рис. 7). Она запускает процессы лигнификации клеточной стенки и отложения каллозы (защитный полисахарид), увеличивает количество активных форм кислорода, активирует синтез защитных PR (Pathogen-Related) белков, фитоалексинов и важных, ответственных за фитоиммунитет гормонов (например, этилена). Все вышеперечисленное доставляет патогенной бактерии немало проблем, поскольку губительно влияет на ее жизнедеятельность. Лигнин и каллоза мешают «непрошенному гостю» проникнуть внутрь растительной клетки, а РR-белки, фитоалексины и активные формы кислорода вполне способны разрушить его структурные компоненты.
Резуховидка Таля — это всем известный арабидопсис, верный помощник биологов и генных инженеров. Чем же он так хорош? Читайте в статье «Модельные организмы: арабидопсис» [16]. — Ред.
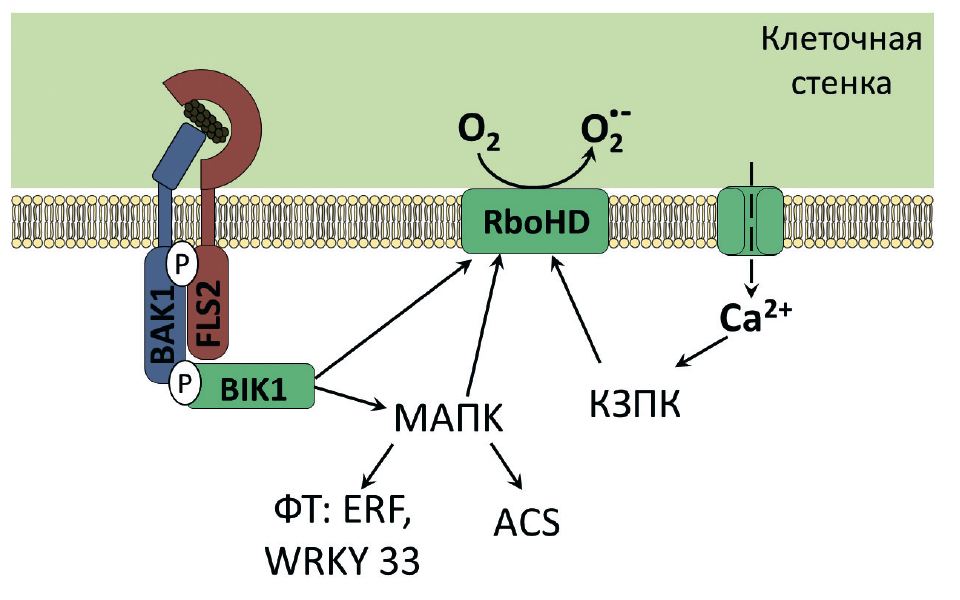
Рисунок 7. Схема развития иммунной реакции растения в ответ на узнавание внеклеточными рецепторами фрагментов бактериального флагеллина.
Рецептор (FLS2) находится на мембране растительной клетки. При проникновении в организм патогенной бактерии он связывается с консервативными участками флагеллина, после чего взаимодействует с двумя ферментами — мембранной киназой BAK1 и цитоплазматической киназой BIK1. В результате киназные домены последних фосфорилируются, что приводит к активации различных флагеллин-индуцируемых ответов. Среди них выброс ионов Ca 2+ в цитоплазму, что активирует кальций-зависимые протеинкиназы (КЗПК), запускающие НАДФН-оксидазу RboHD. НАДФН-оксидаза RboHD образует большое количество свободных форм кислорода — происходит мощный окислительный взрыв, приводящий к гибели клетки фитопатогена. Еще одним результатом фосфорилирования киназного домена BIK1 является запуск МAPK сигнального каскада (МАПК), который активирует ферменты биосинтеза этилена (ASC), НАДФН-оксидазу RboHD, о которой уже шла речь выше, и факторы транскрипции (ФТ) ERF и WRKY33, контролирующие экспрессию защитных генов [9], [17].
Однако, как любой клеточный рецептор, рецептор количественной устойчивости не может быть активирован вечно. Клетке обязательно необходимо его выключить спустя какое-то время, иначе это приведет к истощению ресурсов растения. Поэтому существуют системы, ответственные за разрушение активированных рецепторов, например, убиквитин-лигазная система, которая метит целевые белки особыми убиквитиновыми последовательностями. Эти последовательности распознают протеасомы — белковые комплексы, служащие для разрушения отработанных белков. Другой системой, выключающей рецептор, является механизм эндоцитоза, за счет которого последний оказывается внутри клетки на эндоцитозном пузырьке и не может больше взаимодействовать со своими экстраклеточными лигандами.
Сигнализация количественной устойчивости устроена таким образом, чтобы обеспечить растению эффективную мобилизацию и своевременную демобилизацию ее защитных систем, что позволяет вовремя препятствовать проникновению патогенного организма, не приводя при этом к большим энергетическим затратам.
К сожалению, несмотря на то, что механизмы количественной устойчивости запускаются в ответ на жизненно важные метаболиты фитопатогенов, они далеко не всегда способны защитить растение. Дело в том, что в ходе эволюции патогены выработали механизмы защиты и обмана данной иммунной реакции организма-хозяина. Например, у бактерий есть экзополисахариды, способные в какой-то степени экранировать их консервативные антигенные детерминанты. Также фитопатогены имеют особые системы секреции, позволяющие им доставлять свои специфичные метаболиты, необходимые для блокировки количественной устойчивости, непосредственно внутрь клетки растения.
Качественная (эффектор-индуцированная или вертикальная) устойчивость является более новым типом устойчивости и по характеру эволюции напоминает гонку вооружений. Немного позже объясним, почему.
Качественная устойчивость на то и качественная, что она либо запускается и убивает патоген на все 100%, либо не запускается совсем. В отличие от количественной устойчивости, качественная — высокоспецифична, вплоть до того, что может сработать на один штамм фитопатогена, но совершенно никак не отреагировать на другой. Она запускается в ответ на особые белки фитопатогенов, называемых специфичными эффекторами, уникальными для каждого вида или даже штамма, которые, как правило, секретируются паразитами непосредственно внутрь клетки растения с целью подавления иммунной защиты хозяина, а также для изменения его метаболизма в своих целях. Растения же «научились» распознавать подобные эффекторы патогенов как сигнал опасности и активировать в ответ на них губительные для микроорганизма защитные реакции. Поскольку специфичные элиситоры доставляются преимущественно внутрь клетки растения, то и рецепторы качественной устойчивости (R-белки) расположены в основном в цитоплазме или же в нуклеоплазме, хлоропластах и эндомембранных структурах [9].
Взаимоотношения растения и фитопатогена при данном типе устойчивости характеризуют еще как отношение «ген на ген». Эта теория была разработана Генри Гарольдом Флором в ходе изучения взаимодействия льна (Linum usitatissimum) и ржавчинных грибов (Melampsora lini). Ее основная идея заключается в следующем: иммунитет развивается, когда продукт некоего гена устойчивости растения (вышеупомянутые R-белки) взаимодействует с продуктом гена авирулентности фитопатогена (тот самый специфичный эффектор). Если же продукт гена авирулентости не соответствует продукту гена устойчивости, то развивается заболевание.
Поскольку качественная устойчивость вызывает абсолютную гибель фитопатогена, она оказывает селективное влияние на его популяцию, что приводит к возникновению устойчивых штаммов, секретирующих новые элиситоры, которые организм хозяина не способен распознать. В ответ на это спустя какое-то время у растений появляются дополнительные системы, позволяющие им бороться с устойчивыми патогенами, что запускает очередной цикл естественного отбора у микроорганизмов. Поэтому эволюционное развитие качественной устойчивости у растений часто сравнивают с гонкой вооружений и описывают так называемой моделью «зигзаг» (рис. 8) [18].
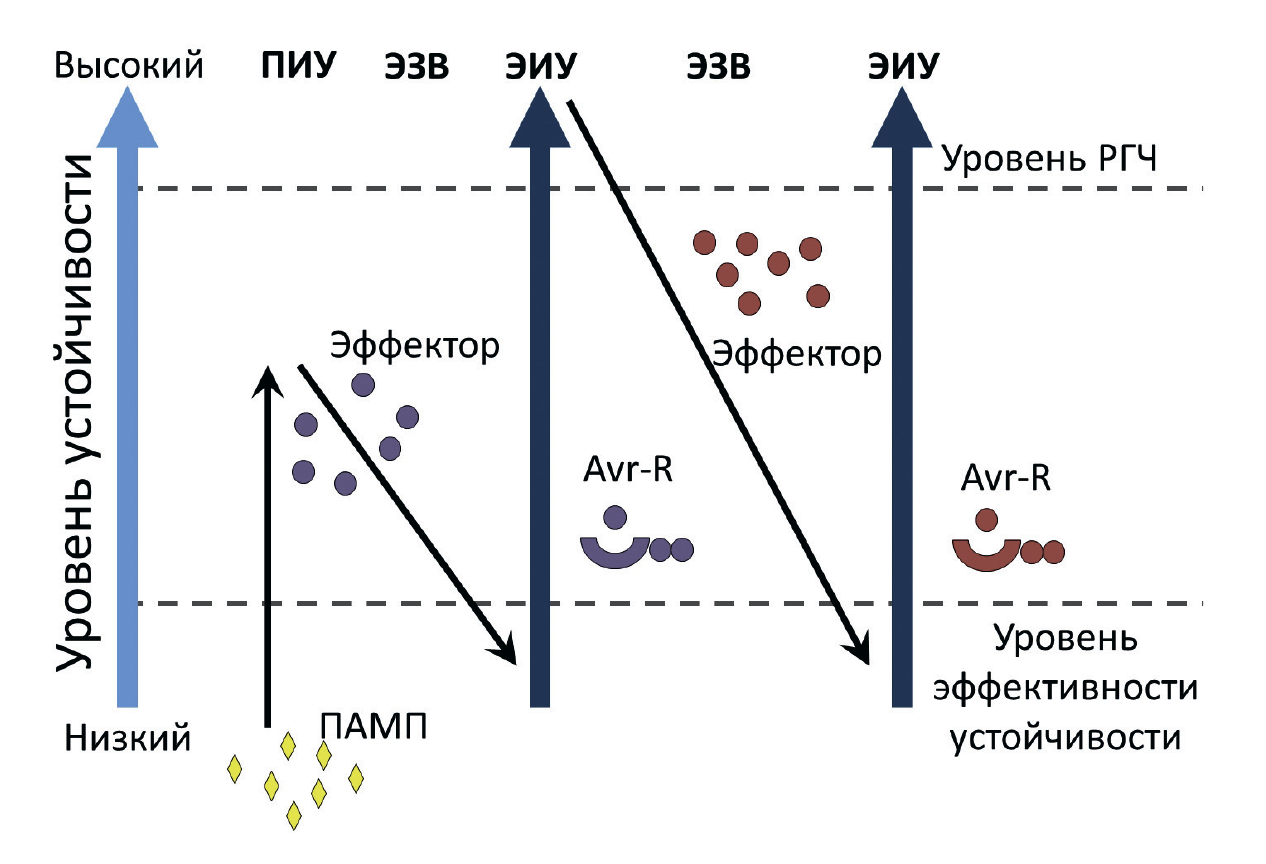
Рисунок 8. Модель «зигзаг», демонстрирующая механизм эволюции качественной устойчивости растений.
Растение успешно распознает патоген-ассоциированный молекулярный паттерн (ПАМП), что приводит к развитию ПАМП-индуцированной устойчивости (ПИУ). Затем у патогена появляется эффектор, способный заблокировать формирование количественного иммунитета, что становится причиной эффектор-зависимой восприимчивости (ЭЗВ). Растительные R-белки распознают эффекторы, блокирующие ПИУ, и активируют эффектор-индуцированную (качественную) устойчивость (ЭИУ). Появление у фитопатогенов в дальнейшем новых эффекторов сводит на нет развитие качественного иммунитета, что опять приводит к развитию заболевания. Появление новых R-белков у растения восстанавливает высокий уровень качественной устойчивости. Уровень РГЧ — это уровень, при котором развивается реакция гиперчувствительности.
Как работают R-белки — рецепторы качественной устойчивости?
R-белки имеют несколько доменов: вариабельный домен, ответственный за распознавание сигнала, нуклеотид-связывающий домен и лейцин-богатую область на С-конце. Когда рецептор связывается с эффектором, изменяется конформация R-белка, и в его нуклеотид-связывающем сайте происходит замена АДФ на АТФ. Последующий гидролиз АТФ обеспечивает взаимодействие активированного рецептора со следующими компонентами каскада защитных реакций.
R-белки могут взаимодействовать с эффекторами напрямую, что отражает концепцию взаимоотношения «ген на ген». Однако зачастую элиситоры связываются со своими рецепторами через специальные «сторожевые белки», которые могут выполнять несколько функций:
- инактивация R-белков в отсутствие фитопатогена;
- участие в развитии горизонтальной устойчивости (являются компонентами внутриклеточного сигнального пути, запускающего количественный иммунитет).
Работает эта «система» следующим образом. Если фитопатогена нет, R-белок связан со своим сторожевым белком (качественный иммунитет выключен). Когда «непрошеный гость» попадает в организм растения, он секретирует внутрь клетки хозяина эффекторы, которые направлены на блокировку сторожевых белков, поскольку те, помимо инактивации рецепторов качественной устойчивости, являются промежуточными звеньями в ПАМП-индуцированном сигнальном каскаде. В попытке выключить развитие количественной устойчивости элиситор фитопатогена связывается со сторожевым белком, что приводит к отсоединению последнего от R-рецептора и активации механизмов качественной устойчивости.
У сторожевых белков есть также эволюционные “родственники” – “белки-приманки”, которые утратили свою функцию в формировании количественной устойчивости, но зато способны связываться с эффекторами фитопатогенов и активировать развитие качественного иммунного ответа по механизму, описанному выше для сторожевых белков.
Подобный подход крайне удобен для растений, поскольку позволяет отслеживать не непосредственно эффекторы фитопатогенов, которые могут быть самыми разнообразными, а состояние мишеней этих элиситоров (сторожевых белков и белков приманок) в растительном организме, что позволяет обеспечить более эффективную и быструю защиту [9].
Физиологически качественная устойчивость может проявляться примерно так же, как и количественная: кальций входит в цитоплазму через открытые ионные каналы; начинают работать Са 2+ -зависимые протеинкиназы, происходит формирование активных форм кислорода, запускается процесс лигнификации РКС, отложения каллозы, синтез PR-белков, фитоалексинов, этилена, салициловой и жасмоновой кислот и т.д. Однако характерной особенностью именно эффектор-индуцированной устойчивости является так называемая реакция гиперчувствительности — особая форма программируемой клеточной смерти (в месте проникновения фитопатогена происходит мощный «окислительный взрыв», приводящий к гибели клеток растения вместе с паразитом). Еще одним отличием является то, что качественный иммунный ответ во много раз сильнее и энергозатратней количественного и запускается, как было описано выше, по совершенно другим молекулярным механизмам [9].
Системная устойчивость
Системные ответы развиваются в незараженных клетках инфицированного растения и позволяют последнему формировать резистентность при повторной инвазии фитопатогена. Существуют индуцированная системная устойчивость (ИСУ) и системная приобретенная устойчивость (СПУ).
Индуцированная системная устойчивость формируется в ответ на различные повреждения (раневые сигналы), вызванные, например, некротрофами или насекомыми, а также на элиситоры PGPR (Plant-Growth Promoting Rhizobacteria) — ризосферных бактерий, которые могут не только стимулировать рост растения, но и способствовать его защите от фитопатогенных микроорганизмов. Подобные элиситоры по химической природе могут быть крайне разнообразными (сидерофоры, липополисахариды, фрагменты жгутиков). Основными сигнальными молекулами ИСУ являются жасмонат и этилен, которые приводят растение в некое состояние готовности и могут активировать синтез защитных PR-белков, фитоалексинов и окислительных ферментов (пероксидаз, липооксигеназ) и т.д. [19].
Системная приобретенная устойчивость является следствием локальной эффектор-индуцированной устойчивости, возникающей в ответ на внедрение фитопатогена, и представляет собой что-то наподобие иммунной памяти. В качестве своеобразного сигнала SOS, от места инфекции по организму растения распространяется целый комплекс различных веществ (азелаиновая кислота, глицеральдегид-3-фосфат и т.д.), который запускает образование салициловой кислоты в клетках, не подверженных атаке фитопатогена, и активацию синтеза защитных веществ (например, PR-белков). Системная приобретенная устойчивость может передаваться по наследству, поскольку в ходе иммунного ответа происходят эпигенетические изменения ДНК растения [9], [19].
Что синтезирует растение в ответ на инвазию патогенов?
Факторы индуцированной устойчивости
Выше мы уже обсудили, какие виды индуцированной устойчивости бывают у растений, какие молекулы распознают растения, где располагаются рецепторы, ответственные за активацию тех или иных сигнальных путей, однако сейчас хотелось бы подробнее остановиться на том, что именно синтезирует растение в ответ на инвазию патогенов.
Читатель мог заметить, что физиологические проявления различных видов индуцированной устойчивости во многом схожи, что говорит о том, что у растения есть определенный набор инструментов, которые оно использует в борьбе с фитопатогенами.
Первое, о чем мы уже упоминали, — это синтез лигнина (полиароматического соединения) в ответ на появление «непрошеного гостя». Лигнин в целом способствует укреплению растительной клеточной стенки, защищает ее структурные углеводы от действия литических ферментов, ограничивает приток воды и питательных веществ к месту расположения фитопатогена. Помимо лигнина, в ответ на инвазию паразита может синтезироваться еще и суберин (глицерид феллоновой [СН3(СН2)19СН(ОН)СООН] и пробковой [СООН(СН2)6СООН] кислот), который откладывается в сосудах и препятствует распространению фитопатогена по растению [11].
Еще одним вариантом иммунного ответа, связанного с укреплением внешних барьеров клетки, является образование полисахарида каллозы, необходимого для экстренной заделки дыр, образовавшихся в растительной клеточной стенке. Каллоза откладывается в виде особых структур — паппил между цитоплазматической мембраной и РКС (рис. 9). Данный полисахарид во многом удобен потому, что его синтез происходит достаточно быстро, и при этом он обладает свойством полупроницаемости — подобно бывшей на этом месте растительной клеточной стенке [20].
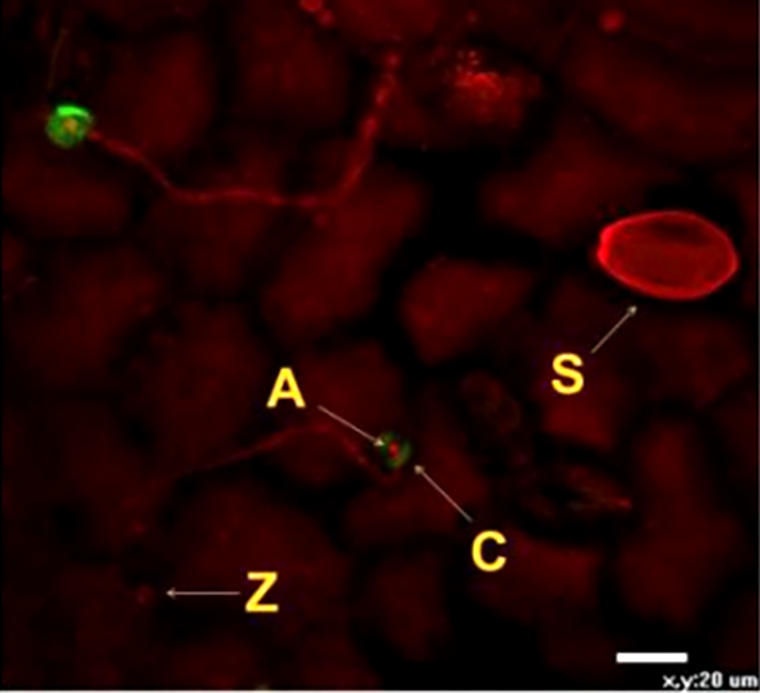
Рисунок 9. Формирование паппил из каллозы клетками огурца. Паппилы (C) образуются вокруг аппрессории фитопатогенного гриба Pseudoperonospora cubensis (A), которая образовалась на конце зародышевой трубки, выросшей из зооспоры (Z). Буквой S обозначен спорангий гриба. Процесс образования каллозы часто происходит и при бактериальной инвазии.
Помимо непосредственного усиления барьерных функций РКС, растения синтезируют множество защитных веществ. Что такое фитоалексины с химической точки зрения, мы уже подробно говорили в одной из предыдущих глав, поэтому теперь обратим внимание на другие метаболиты.
Патогенез-ассоциированные белки (Pathogenesis-related proteins, PR) являются факторами как локальной, так и системной индуцированной устойчивости. Существует множество групп PR-белков; при этом все еще неизвестна роль многих из них в защите растения. Среди PR-белков, чьи функции известны, встречаются ферменты (глюканазы и хитиназы), расположенные либо в апопласте, либо в вакуоли, участвующие в деградации клеточных стенок фитопатогенов, пермантины, дефензины и тионины — белки, нарушающие работу мембранных каналов и тем самым приводящие к лизису клеток, ингибиторы различных микробных гидролаз и многие другие [21].
Также растения способны синтезировать в ответ на инвазию бактериальных патогенов так называемые репрессоры чувства кворума. Что это такое?
«Чувство кворума» — это особая регуляторная система, распространенная среди прокариот. По сути, это некий феномен межклеточной коммуникации у бактерий с помощью низкомолекулярных соединений, позволяющий микроорганизмам действовать согласованно при достижении определенного порога плотности популяции. На примере фитопатогенов этот феномен можно продемонстрировать следующим образом. Когда бактерия проникает в организм растения, она прекрасно «понимает», что, если она сейчас начнет синтезировать ферменты, разрушающие клеточную стенку хозяина, то последний явно этому не обрадуется и уничтожит ее в ближайшее время. Однако, если она подождет, размножится и только после этого начнет синтезировать те же гидролазы, то растению будет намного сложнее справиться с инфекцией.
Но что же при этом происходит на биохимическом уровне? Дело в том, что низкомолекулярные соединения, служащие медиаторами чувства кворума, при достижении бактериями определенной плотности активируют синтез самих себя по принципу положительной обратной связи, а также синтез ферментов, участвующих в развитии бактериальной инфекции (рис. 10).
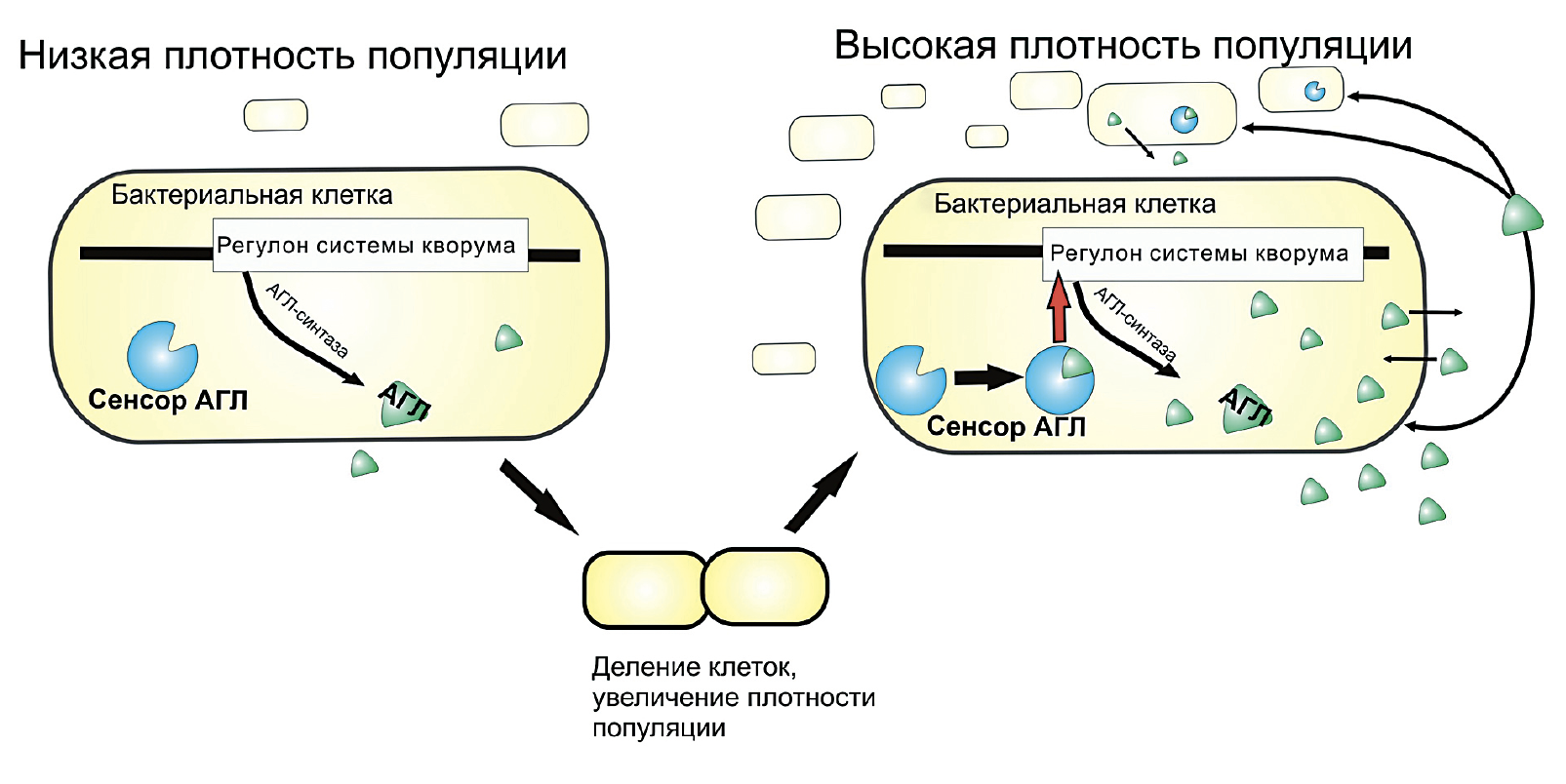
Рисунок 10. Схема регуляторной системы чувства кворума бактерий, где в качестве медиатора выступает ацил-гомосерин лактон (АГЛ). Когда плотность популяции низкая, АГЛ вырабатывается мало, следовательно, экспрессия генов факторов вирулентности затруднена. При увеличении плотности популяции бактерий данного медиатора становится много, и тогда он связывается со своим внутриклеточным сенсором. Комплекс «Сенсор + АГЛ» активирует экспрессию соответствующих генов (регулон системы кворума), кодирующих, например, литические ферменты, разрушающие растительную клеточную стенку, а также ген АГЛ-синтазы, тем самым способствуя образованию ацил-гомосерин лактона.
Что же делают растительные репрессоры чувства кворума? По своей структуре они похожи на низкомолекулярные медиаторы системы кворума бактерий, поэтому способны связываться с внутриклеточными рецепторами этих соединений у микроорганизмов и подавлять экспрессию соответствующих генов фитопатогенов. Иногда растения поступают еще хитрее: они «привлекают» в свою ризосферу такие микроорганизмы (например, представителей рода Bacillus и Rhodococcus), которые способны разрушать медиаторы системы чувства кворума фитопатогенных бактерий [9], [22].
Еще одним часто упоминавшимся выше вариантом ответа растения на внедрение паразитов является синтез активных форм кислорода. Наверное, ни для кого не секрет, что активные формы кислорода (гидроксил-радикал, перекись водорода, супероксид анион и другие) крайне опасны как для растительной, так и для животной клетки, поскольку способны активировать цепные реакции окисления различных биологических молекул от липидов до ДНК. Так при чем тут защита от фитопатогенов? Увеличение синтеза АФК характерно для количественной устойчивости. Активные формы губительно действуют на клетки фитопатогенов, способствуют сшивке полимеров растительных клеточных стенок, являются компонентами защитных сигнальных каскадов. Также они принимают участие в развитии реакции гиперчувствительности (РГЧ) — особого типа быстрой программируемой клеточной смерти (качественная устойчивость). Она приводит к полному уничтожению фитопатогена вместе с клеткой растения-хозяина и является своего рода экстренной мерой. В данном случае активный синтез АФК осуществляется в основном за счет активации НАДФ-оксидазы, расположенной в плазмалемме. Этот фермент окисляет НАДФ, находящийся в цитоплазме, и переносит электрон на кислород, расположенный в апопласте, приводя к формированию оксид радикала. Развитию окислительного взрыва также способствует ингибирование антиоксидантных ферментов, таких как каталаза или пероксидаза, участвующих в детоксикации перекиси водорода. Обязательным регулятором РГЧ является салициловая кислота [23].
Фитогормоны и фитоиммунитет
Мы знаем, что большинство процессов, происходящих в нашем организме, регулируется веществами различной химической природы: гормонами, которые синтезируются специальными органами — железами внутренней секреции. Растения в этом случае не являются исключением. У них нет специализированных тканей, ответственных за синтез подобных веществ, но растительных гормонов существует довольно много. Это и классические гормоны: ауксины, гиббереллины, цитокины — их еще называют ростовыми, а также этилен, абсцизины (абсцизовая кислота и ее производные) [24].
Фитогормоны играют огромную роль в формировании иммунного ответа — от них во многом зависит степень устойчивости или восприимчивости растения к конкретному патогену. Более того, иногда патогенные организмы способны влиять на гормональный статус растения, заставляя его работать так, как им выгодно.
Основными гормонами, которые играют главную роль в фитоиммунитете, считаются салициловая, жасмоновая кислоты и их производные, а также этилен.
Салициловая кислота
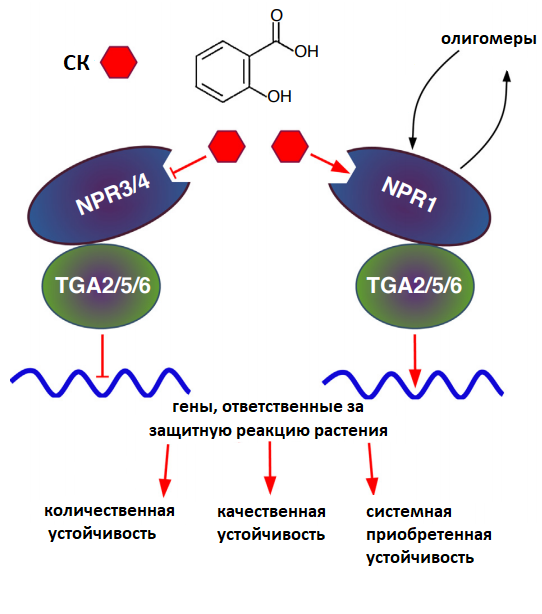
Рисунок 11. Схема сигнального пути салициловой кислоты внутри клетки растения. СК участвует в развитии как качественной (эффектор-индуцированной устойчивости), так и количественной (паттерн-индуцированной устойчивости) и системной приобретенной устойчивости. У СК существует 3 основных внутриклеточных рецептора — NPR1, NPR 3, NPR 4. В отсутствии фитогормона белки NPR3/4 ингибируют активность факторов транскрипции TGA2/5/6, ответственных за экспрессию генов, участвующих в реализации различных видов иммунного ответа растений. СК ингибирует действие этих белков, следовательно TGA2/5/6 могут быть активированы под действием второго рецептора фитогормона — NPR1, который после связывания с СК переходит из мультимерного состояния в олигомерное и способен активировать факторы транскрипции.
Первое сообщение о том, что салициловая кислота (СК) играет роль в защите растений от фитопатогенов, относится к 1979 году. Тогда было показано, что обработка табака СК способствует повышению его устойчивости к вирусу табачной мозаики. Позднее ученые установили, что салициловая кислота регулирует множество иммуномодулирующих процессов, принимает участие в развитии локальной количественной и качественной, а также системной приобретенной устойчивости (рис. 11).
Салициловая кислота может ингибировать каталазу и аскорбат пероксидазу — ферменты, участвующие в детоксикации перекиси водорода, вызывая тем самым в растении окислительный взрыв, что, в свою очередь, приводит к гибели фитопатогена вместе с клетками организма-хозяина (реакция гиперчувствительности).
Стоит отметить, что количество салициловой кислоты, задействованное при количественной, системной приобретенной и качественной устойчивости, достаточно разнится. Дело в том, что в первых двух вариантах синтез СК активизируется на значительно более низком уровне по сравнению с последним, что определяет в том числе и силу иммунного ответа. В случае системной приобретенной устойчивости салициловая кислота служит в основном в качестве сигнала, оптимизирующего транскрипцию защитных генов (PR-белков и др.).
К слову, стоит сказать, что увеличение количества СК далеко не всегда приводит к усилению защиты растения от паразитов. Для некоторых фитопатогенов (например, Xanthomonas campestris pv. vesicatoria), наоборот, характерно накопление данного фитогормона при развитии инфекции, а снижение его количества приводит к повышению устойчивости растения. Подобная картина характерна и для некротрофов, которым выгодна смерть растительных клеток под действием салициловой кислоты.
Таким образом, данный фитогормон может рассматриваться не только как фактор устойчивости, но и как фактор восприимчивости — все будет зависеть как от патогена, так и от степени активации СК-индуцируемого ответа [9], [26].
Жасмоновая кислота и этилен
Если мы говорим о защите растений от фитопатогенов, жасмоновая кислота (ЖК) и этилен обычно работают вместе. ЖК служит ключевым компонентом защиты от насекомых-фитофагов, но при этом она также играет важную роль во взаимодействии растений и микроорганизмов. ЖК — это продукт перекисного окисления жирных кислот. Рецепторами жасмоновой кислоты в клетке являются белок COI1 — компонент комплекса, участвующего в деградации белков, и белки JAZ, которые связываются с фактором регуляции транскрипции MYC2 и препятствуют экспрессии ЖК-индуцируемых генов. Жасмоновая кислота в этом случае выступает своего рода клеем между белками COI1 и JAZ, что в конечном итоге приводит к деградации JAZ-белков и активации экспрессии ЖК-индуцируемых генов. ЖК также активирует экспрессию генов, кодирующих ферменты ее собственного биосинтеза и гены JAZ-белков, что обеспечивает регуляцию сигнала, вызываемого этим фитогормоном по принципу положительной и отрицательной обратной связи [27].
Что же касается этилена, то его роль в растении воистину многогранна. Этот фитогормон регулирует множество физиологических процессов в растении: созревание плодов, старение, реакцию на абиотический стресс и т.д [24]. В растениях этилен синтезируется из 1‑аминоциклопропан-1‑карбоновой кислоты, которая является промежуточным продуктом метаболизма метионина. В рецепции данного фитогормона участвует множество белков: ETR1, ETR2, EIN4, ERS1, ERS2, расположенных на мембранах эндоплазматического ретикулума или аппарата Гольджи. Этилен является ключевым регулятором образования в сосудах ксилемы тилл (особых выростов клеток осевой или лучевой паренхимы, заполняющих полости сосудов и трахеид в поврежденных участках), которые блокируют водный транспорт, что может иметь двойной эффект (рис. 12) [28].
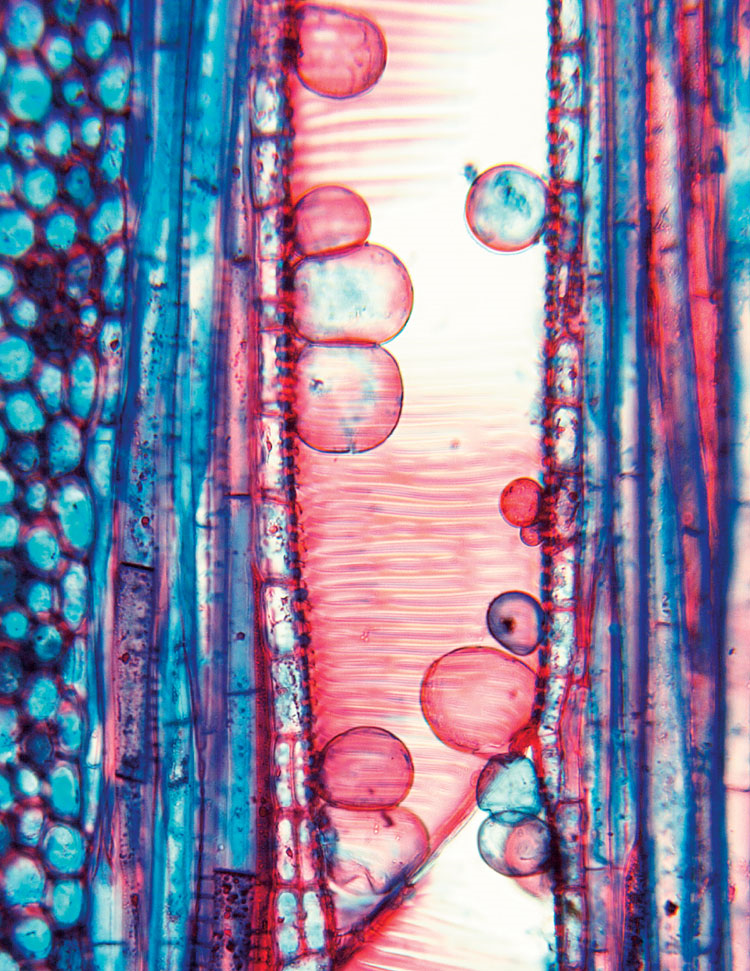
Рисунок 12. Ксилема растения с тиллами — пузыревидными выростами соседних паренхимных клеток, которые выпячиваются непосредственно в просвет сосуда.
Таким образом, с одной стороны, растение препятствует распространению фитопатогена, но с другой стороны, для бактерий это может оказаться выгодным, поскольку образование тилл снижает ксилемный ток в растении, что способствует активной коммуникации бактерий между собой.
В целом, работая вместе, жасмоновая кислота и этилен активируют синтез фитоалексинов, PR-белков, а также стимулируют лигнификацию клеток растения [29]. Этилен и ЖК обеспечивают активацию систем количественной и индуцированной системной устойчивости. Учитывая тот факт, что салицил-индуцируемая и жасмонат-индуцируемая сигнальные системы являются взаимоисключающими, некоторые микроорганизмы «специально» активируют ЖК/этилен-опосредуемые ответы, поскольку те могут способствовать развитию инфекции, вместо, например, салицил-индуцируемых.
А зачем нам изучать, как работает иммунитет растений?
Как видно из вышесказанного, иммунная система растений крайне запутанная, интересная и во многом еще плохо изученная область, которая имеет огромный потенциал в плане практического применения фундаментальных знаний.
Но зачем человеку изучать работу целой иммунной системы растений, если против бактерий, грибов и других убийц урожая давно придумано идеальное оружие — химические «яды»? Эх, если бы все было так просто! Во-первых, пестициды — крайне опасные вещества: они могут обладать канцерогенными и мутагенными свойствами, — а потому их применение и содержание в растениях должно жестко контролироваться, что происходит далеко не всегда. Более того, загрязнение окружающей среды пестицидами является серьезной экологической проблемой, а следовательно, применение таких веществ не может быть лучшим решением [30].
Подробнее о том, какими бывают пестициды и как осуществляется контроль за ними в России и в мире, можно прочесть в спецпроекте «Пестициды» на «Биомолекуле». — Ред.
Во-вторых, если мы постоянно будем «бить фитопатоген дубинкой по голове», то мы лишь поспособствуем появлению новых, устойчивых к дубинке штаммов. Да-да, здесь работает то же правило, что и с бактериальной антибиотикорезистентностью!
Другой вариант — создавать сорта растений, абсолютно устойчивых к фитопатогенам, например, методом генной инженерии . Но и здесь есть несколько подвохов. Во-первых, подобные сорта действуют на патоген так же, как пестициды. В геном таких растений часто встраивают некий ген устойчивости, продукт которого прямо или косвенно убивает микроорганизм, что в целом оказывает тот же самый селективный эффект на популяцию фитопатогенов, что и применение токсичных химических веществ. Такими генами устойчивости могут быть последовательности, кодирующие ферменты, непосредственно действующие на патогены (трансгенный картофель, экспрессирующий ген лизоцима бактериофага Т4 устойчив к Erwinia carotovora atroseptica); гены, чьи продукты ингибируют апоптоз клеток растения (синтез в табаке белков CED-9 нематоды и Bcl-2 человека значительно повышает его невосприимчивость к грибу-некротрофу Sclerotinia sclerotiorum) или, наоборот, стимулируют гибель клеток, вызывая тем самым реакцию гиперчувствительности [31], [32]. Еще одним вариантом может быть искусственное усиление экспрессии уже имеющихся у растений генов, кодирующих защитные белки. Во-вторых, значимой проблемой генетически измененных растений является трудность создания сортов, которые были бы устойчивы сразу к нескольким фитопатогенам.
Интересной альтернативой перечисленным способам борьбы с болезнями растений стало подавление, сдерживание патогенов в рамках латентных инфекций — таких, при которых микроорганизм может размножаться в организме растения, но не вызывать в нем патологических процессов. Читатель вправе спросить: «А возможен ли такой расклад в природе»? Да, так бывает, и довольно часто. Латентные инфекции — результат длительной совместной эволюции патогена и хозяина, но результат не окончательный. Ведь от любви до ненависти один шаг. Пока остается не ясным, что заставляет микроорганизм быть добрым или агрессивным соседом, учитывая то, что в любой момент он может перейти от мирных действий к атаке — и это следствие недостатка наших фундаментальных знаний [9].
Но изучение растительно-микробных сообществ уже дает свои плоды. Например, было выявлено, что подселение к растению «полезной микрофлоры» способно увеличить его устойчивость к фитопатогенным микроорганизмам . Симбиотические ризобактерии, как уже было сказано выше, способны запускать в растении жасмонат-индуцируемые иммунные ответы, конкурировать с фитопатогенами за важные микроэлементы (например, железо), а также синтезировать антибиотики, пагубно влияющие на патогенную микрофлору. Поэтому обработка сельскохозяйственных культур пробиотиками (смесью полезных микроорганизмов) является перспективным направлением в агропромышленности, не лишенным, правда, своих недостатков (например, после высадки в поле растения с симбиотическими бактериями вся его полезная микрофлора может быть вытеснена естественными штаммами).
Подробнее об использовании пробиотиков для борьбы с фитопатогенами читайте в статье «Пробиотики для растений: как накормить растущий мир» [34]. — Ред.
Таким образом, изучение фитоиммунитета, а также растительно-микробных сообществ в целом может подарить нам новые возможности в борьбе с заболеваниями сельскохозяйственных культур.
Основные типы болезней растений и их проявление в зависимости от уровня паразитизма возбудителя
Ложная мучнистая роса (пероноспороз)
 Ложная мучнистая роса, или пероноспороз больше распространяется в зонах достаточного увлажнения, особенно в годы с прохладным летом. Поражаются листья, меньше стебли и другие надземные органы. На листьях с верхней стороны появляются желтые пятна, а с нижней — густой, пушистый грязно-фиолетовый налет.
Ложная мучнистая роса, или пероноспороз больше распространяется в зонах достаточного увлажнения, особенно в годы с прохладным летом. Поражаются листья, меньше стебли и другие надземные органы. На листьях с верхней стороны появляются желтые пятна, а с нижней — густой, пушистый грязно-фиолетовый налет.
Возбудитель болезни — низший гриб Peronospora viciaesativae Gaeum. Налет, который появляется на нижней стороне листьев, является его конидиальным спороношением. Конидиеносцы выступают из устьиц пучками (по 2-6) с 4-9-кратным дихотомическим ветвлением. Конидии одноклеточные, эллиптические, желтоватые, размером 9-27 х 8-24 мкм.
Кроме конидиального спороношения, гриб в пораженных тканях образует желтые сетчатые ооспоры диаметром 29-46 мкм. Зимует возбудитель на остатках растений в виде ооспор.
При интенсивном развитии заболевания листья буреют, засыхают и опадают, а зерно в бобах недоразвивается.
Ржавчина  Обнаруживается в течение всей вегетации растений. Вначале на молодых листьях появляются мелкие желтые эции, а позже светло-коричневые урединиопустулы. К концу вегетации растений на стеблях и листьях образуются темно-коричневые или черные телиопустулы. Пораженные листья усыхают и опадают. Нередко поражение в виде урединио- и телиопустул обнаруживается и на бобах.
Обнаруживается в течение всей вегетации растений. Вначале на молодых листьях появляются мелкие желтые эции, а позже светло-коричневые урединиопустулы. К концу вегетации растений на стеблях и листьях образуются темно-коричневые или черные телиопустулы. Пораженные листья усыхают и опадают. Нередко поражение в виде урединио- и телиопустул обнаруживается и на бобах.
Возбудитель болезни — базидиальный гриб Uromyces fabae d By.
Спермогонии и эции формируются на нижней стороне листьев небольшими группами, часто концентрическими кругами. Эциоспоры шаровидные или эллиптические, размером 14-21 х 18-26 мкм, с мелкобородавчатой оболочкой.
Урединиопустулы размером до 1,5 мм, образуются с обеих сторон листа. Урединио-споры одноклеточные, шаровидные или яйцевидные, 17-26 х 21-30 мкм, желтоватые, с оболочкой, покрытой редкими шипиками.
Телиопустулы также формируются с обеих сторон листа. Телиоспоры 25-40 х 18-29 мкм, на желтоватой ножке, одноклеточные, округлые или яйцевидные, к основанию суженные, часто на вершине сосочкообразно вытянутые, с гладкой бурой оболочкой.
Гриб зимует на остатках растений в виде телиоспор, которые весной, прорастая, образуют базидии с базидиоспорами. От них и происходит первичное заражение растений.
При интенсивном развитии болезни недобор урожая зеленой массы и семян вики может достигать 20-30%.
Аскохитоз
 Проявляется на листьях, стеблях и бобах в виде неотчетливых, сливающихся пятен с приплюснутыми черными пикнидами, расположенными неправильными концентрическими кругами.
Проявляется на листьях, стеблях и бобах в виде неотчетливых, сливающихся пятен с приплюснутыми черными пикнидами, расположенными неправильными концентрическими кругами.
Возбудитель болезни — несовершенный гриб Ascochyta punctata Naumov. Его пикниды размером 180-130 мкм, пикно-споры двухклеточные, бесцветные, цилиндрические, с закругленными концами, размером 18 х 4,5 мкм.
При интенсивном развитии болезни листья преждевременно опадают, пораженные стебли надламываются. При поражении бобов грибница возбудителя проникает в семена. Сохраняется гриб в семенах и остатках растений.
Септориоз
 Поражает все надземные органы растений. На листьях появляются беловатые пятна с темнокоричневой каймой. Со временем на них образуются черные точковидные пикниды чечевицеобразной формы.
Поражает все надземные органы растений. На листьях появляются беловатые пятна с темнокоричневой каймой. Со временем на них образуются черные точковидные пикниды чечевицеобразной формы.
Возбудитель болезни — несовершенный гриб Septoria viciae West. В его пикнидах образуются тонкие прямые одноклеточные бесцветные пикноспоры размером 30-60 х 2,5 мкм.
Зимует гриб на остатках пораженных растений, а иногда и на семенах в виде грибницы или пикнид с пикноспорами. Заболевание приводит к недобору зеленой массы и семян.
Мозаика
 Вызывает характерное изменение окраски и курчавость листьев, деформацию бобов, а иногда и некротические полосы на листьях и стеблях.
Вызывает характерное изменение окраски и курчавость листьев, деформацию бобов, а иногда и некротические полосы на листьях и стеблях.
Возбудитель болезни — Pisum virus 2 Smith. Поражает он также горох, кормовые бобы, клевер, люпин узколистный, донник и другие бобовые растения.
Переносчиками вируса являются тли. Болезнь не вызывает гибели растений, но может быть причиной значительного недобора урожая зеленой массы и семян.
Болезни горох
- Печать
Бактериоз
 На листьях, стеблях и бобах появляются сравнительно крупные, неправильной или округлой формы коричневые с маслянистой каймой пятна, которые иногда сливаются. Болезнь вызывают бактерии Pseudomonas pisi Sackett.
На листьях, стеблях и бобах появляются сравнительно крупные, неправильной или округлой формы коричневые с маслянистой каймой пятна, которые иногда сливаются. Болезнь вызывают бактерии Pseudomonas pisi Sackett.
Растения заражаются через устьица и ранки. Бактерии распространяются по межклетникам, но нередко через сосудистую систему попадают в бобы и поражают семена, вследствие чего на них появляются вдавленные пятна.
Источником инфекции являются зараженные семена и неперегнившие остатки растений в почве.
Относительно устойчивы к бактериозу сорта Уладовский 10, Уладовский юбилейный и Рамонский 77.
Бледный аскохитоз
 Проявляется в виде светло-каштановых пятен с темно-коричневой каймой на бобах, реже на листьях и стеблях. На бобах и листьях пятна округлые, диаметром до 9 мм, а на стеблях и черешках — удлиненные. В центре пятен образуются темно-коричневые пикниды. Иногда при поражении растений в конце вегетации на бобах и стеблях пятна не образуются, но появляются многочисленные пикниды, покрывающие их поверхность. Пораженные семена морщинистые, со светло-желтыми слабо выраженными пятнами.
Проявляется в виде светло-каштановых пятен с темно-коричневой каймой на бобах, реже на листьях и стеблях. На бобах и листьях пятна округлые, диаметром до 9 мм, а на стеблях и черешках — удлиненные. В центре пятен образуются темно-коричневые пикниды. Иногда при поражении растений в конце вегетации на бобах и стеблях пятна не образуются, но появляются многочисленные пикниды, покрывающие их поверхность. Пораженные семена морщинистые, со светло-желтыми слабо выраженными пятнами.
Возбудитель бледного аскохитоза — гриб Ascochyta pisi Libert. Поражает он только горох, на котором образуется бесполое спороношение — пикниды с пикноспорами. Пикниды шаровидные, слегка приплюснутые, 200-212 мкм в диаметре. Пикноспоры продолговатые, с закругленными концами, с одной, редко двумя или тремя перегородками, размером 9,6-19 X 8,5-6 мкм. Прорастают пикноспоры в капельной влаге. Температурный оптимум заражения 18-20°.
Гороховая цистообразующая нематода

Heterodera goettingiana Liebscher. Поражает горох, вику, чину, чечевицу и люпин. Распространяется очагами. Больные растения отстают в росте и часто имеют карликовый вид. Листья их желтеют, начиная от нижних к верхним, и преждевременно отмирают. Сильно пораженные растения увядают и отмирают полностью. Корни больных растений очень тонкие, вялые и мало ветвятся. На них в весенний период сперва обнаруживаются личинки, затем взрослые нематоды, а в середине лета — белые цисты.
Самки лимонообразные или округлые, размером 0,6-0,8 X 0,5-0,6 мм. Самцы червеобразные, длиной около 1,29 мм. На растениях, зараженных этой нематодой, интенсивно развивается корневая гниль.
Деформирующая мозаика
 Проявляется в виде крапчатости, морщинистости, а также курчавости листьев и прицветников. Сначала на листьях появляются хлоротичные пятна, которые со временем становятся белыми и почти прозрачными.
Проявляется в виде крапчатости, морщинистости, а также курчавости листьев и прицветников. Сначала на листьях появляются хлоротичные пятна, которые со временем становятся белыми и почти прозрачными.
При поражении молодых растений их развитие прекращается, а возле поверхности почвы образуется розетка уродливых листьев. Нередко наблюдается деформация бобов вследствие неравномерного разрастания тканей створок. Семена с пораженных бобов имеют более интенсивную желтую окраску.
Возбудитель болезни — вирус Pisum virus 1 Smith. Кроме гороха, может поражать кормовые бобы, сою и душистый горошек. Переносится тлями. С семенами не распространяется. Меньше поражается им сорт гороха Уладовский юбилейный.
Корневая гниль
 Распространена во всех районах возделывания гороха в период его вегетации. Листья пораженных растений желтеют, скручиваются, засыхают и опадают. Сильно ослабленные растения погибают, а более устойчивые отстают в росте. Над местом поражения главного корня появляется большое количество тонких боковых корешков. Во влажную погоду пораженные места покрываются белым или розоватым налетом с оранжевыми или розоватыми подушечками (спородохии).
Распространена во всех районах возделывания гороха в период его вегетации. Листья пораженных растений желтеют, скручиваются, засыхают и опадают. Сильно ослабленные растения погибают, а более устойчивые отстают в росте. Над местом поражения главного корня появляется большое количество тонких боковых корешков. Во влажную погоду пораженные места покрываются белым или розоватым налетом с оранжевыми или розоватыми подушечками (спородохии).
Иногда наблюдается трахеомикозное увядание. Листья и верхушки стебля поникают и быстро засыхают. Сосуды корней, стеблей, черешков листьев, цветоножек приобретают красно-коричневую с различными оттенками окраску, особенно хорошо заметную на поперечном срезе. Больные растения легко выдергиваются из почвы.
В начале развития у пораженных растений буреет подсемядольное колено, затем коричневатые пятна появляются на прикорневой части стебля, главном корне и корневых волосках, которые постепенно теряют тургор, темнеют и отмирают.
Возбудители заболевания — несовершенные грибы рода Fusarium Link (чаще F. oxysporum Schlecht f. pisi Bilai, F. culmorum Sacc. и др.), а иногда Thielaviopsis basicola Ferr., Rhizoctonia solani Kuehn. Грибы рода Fusarium при благоприятных для них условиях образуют большое количество макро- и микроконидий, что способствует их быстрому распространению.
Макроконидии многоклеточные, серповидные или веретеносерповидные, а микроконидии одноклеточные или с одной перегородкой. Первые из них образуются на слаборазветвленных конидиеносцах или спородохиях, а вторые — на гифах грибницы.
Возбудители болезни обитают в почве, на остатках растений и могут распространяться с семенами, на которых находятся в виде грибницы в семенной оболочке.
Развитию корневой гнили способствуют высокие температуры (среднесуточные 19- 23, максимальные 34-39°) и низкая относительная влажность воздуха (45-60%).
Корневая гниль резко уменьшает адсорбирующую и поглотительную способность корней, а поражение сосудистой системы вызывает ее интоксикацию. Пораженные растения не плодоносят или образуют щуплые семена.
Относительно меньше поражаются сорта Неосыпающийся 1, Уладовский 10, а из овощных сортов Кубанец 1126 (лущильный) и др.
Ложная мучнистая роса (пероноспороз)
 Ложная мучнистая роса, или пероноспороз больше распространяется в зонах достаточного увлажнения, особенно в годы с прохладным летом. Поражаются листья, меньше стебли и другие надземные органы. На листьях с верхней стороны появляются желтые пятна, а с нижней — густой, пушистый грязно-фиолетовый налет.
Ложная мучнистая роса, или пероноспороз больше распространяется в зонах достаточного увлажнения, особенно в годы с прохладным летом. Поражаются листья, меньше стебли и другие надземные органы. На листьях с верхней стороны появляются желтые пятна, а с нижней — густой, пушистый грязно-фиолетовый налет.
Возбудитель болезни — низший гриб Peronospora viciaesativae Gaeum. Налет, который появляется на нижней стороне листьев, является его конидиальным спороношением. Конидиеносцы выступают из устьиц пучками (по 2-6) с 4-9-кратным дихотомическим ветвлением. Конидии одноклеточные, эллиптические, желтоватые, размером 9-27 х 8-24 мкм.
Кроме конидиального спороношения, гриб в пораженных тканях образует желтые сетчатые ооспоры диаметром 29-46 мкм. Зимует возбудитель на остатках растений в виде ооспор.
При интенсивном развитии заболевания листья буреют, засыхают и опадают, а зерно в бобах недоразвивается.
Мучнистая роса
 Распространена в Степи и Лесостепи Украины, а также в Центральной черноземной зоне России. Проявляется в виде белого или мучнистого налета на листьях, преимущественно с верхней стороны, стеблях, прицветниках, цветках и бобах. Со временем налет уплотняется, становится грязно-серым вследствие формирования клейстотециев. При сильном развитии болезни пораженные части растений, приобретают грубую консистенцию и отмирают.
Распространена в Степи и Лесостепи Украины, а также в Центральной черноземной зоне России. Проявляется в виде белого или мучнистого налета на листьях, преимущественно с верхней стороны, стеблях, прицветниках, цветках и бобах. Со временем налет уплотняется, становится грязно-серым вследствие формирования клейстотециев. При сильном развитии болезни пораженные части растений, приобретают грубую консистенцию и отмирают.
Возбудитель болезни — сумчатый гриб Erysiphe communis Grev. f. pisi Dietrich. Его грибница поверхностная, прикрепляется к тканям растений аппрессориями, а внутрь клеток проникают гаустории. На грибнице появляются неразветвленные, слегка вытянутые конидиеносцы, на вершинах которых формируются одноклеточные бесцветные эллиптические конидии размером 28-44 X 17-18 мкм. Они легко переносятся ветром, капельками дождя и насекомыми, вызывая быстрое распространение болезни на посевах. Оптимальными условиями для прорастания конидий являются температура 20° и высокая относительная влажность воздуха.
К концу вегетации растений на грибнице появляются бледно-желтые, затем коричневато-черные клейстотеции. Они шарообразные, диаметром 84-169 мкм. Зимой в них образуется по 5-8 сумок, в каждой из которых находится по 2-6 сумкоспор.
Сумки эллиптические, суженные книзу, неравносторонние, размером 42-67 X 21-30 мкм. Сумкоспоры бесцветные, эллиптические, размером 16-24 X 19-12 мкм. Созревают они весной и служат первичным источником заражения посевов гороха. Попав на молодые растения, сумкоспоры быстро прорастают, образуют один или два ростка, которые развиваются в грибницу. Заболевание интенсивно распространяется при температуре 20-25° и относительной влажности воздуха 70-80%. В этих условиях инкубационный период болезни длится 4-5 дней.
Очень сильно развивается мучнистая роса на поздних посевах гороха. Возбудитель ее зимует в поле на остатках растений в виде клейстотециев, из которых весной распространяются сумкоспоры. Устойчивых к болезни сортов гороха нет. Меньше поражается Рамонский 77.
Обыкновенная мозаика
 Характеризуется вначале посветлением жилок, а затем пожелтением отдельных участков листа. Пораженные листья иногда приобретают удлиненную форму, растения отстают в росте и дают низкий урожай.
Характеризуется вначале посветлением жилок, а затем пожелтением отдельных участков листа. Пораженные листья иногда приобретают удлиненную форму, растения отстают в росте и дают низкий урожай.
Возбудитель болезни — вирус Pisum virus 2 Smith. Сохраняется он на многолетних травах, в частности на клевере, переносится тлями. С семенами не передается. В сухих листьях может сохранять жизнеспособность в течение 45-50 дней.
Оливковая плесень
 Проявляется на листьях, стеблях и бобах в виде оливково-коричневого, почти черного бархатистого налета, который уплотняется и часто покрывает органы растений сплошными пятнами. Особенно интенсивно заболевание развивается в период созревания зерна во влажную погоду.
Проявляется на листьях, стеблях и бобах в виде оливково-коричневого, почти черного бархатистого налета, который уплотняется и часто покрывает органы растений сплошными пятнами. Особенно интенсивно заболевание развивается в период созревания зерна во влажную погоду.
Возбудитель болезни — несовершенный гриб Cladosporium herbarum Fr. Его грибница темного цвета, находится под поверхностью пораженной ткани, на которой снаружи в виде пучков образуются оливко-во-коричневые конидиеносцы с конидиями. Конидиеносцы простые, иногда к вершине немного разветвлены, толщиной 5-10 мкм.
Конидии удлиненные, цилиндрические, вначале без перегородок, а позже с 3-4 перегородками, мелкощетинистые, размером 12-28 X 6-7 мкм, иногда почкуются.
Гриб поражает растения многих видов. Сохраняется на растительных остатках и зерне. Сильно пораженные семена имеют меньшую массу и теряют всхожесть.
Ржавчина
 Обнаруживается в течение всей вегетации растений. Вначале на молодых листьях появляются мелкие желтые эции, а позже светло-коричневые урединиопустулы. К концу вегетации растений на стеблях и листьях образуются темно-коричневые или черные телиопустулы. Пораженные листья усыхают и опадают. Нередко поражение в виде урединио- и телиопустул обнаруживается и на бобах.
Обнаруживается в течение всей вегетации растений. Вначале на молодых листьях появляются мелкие желтые эции, а позже светло-коричневые урединиопустулы. К концу вегетации растений на стеблях и листьях образуются темно-коричневые или черные телиопустулы. Пораженные листья усыхают и опадают. Нередко поражение в виде урединио- и телиопустул обнаруживается и на бобах.
Возбудитель болезни — базидиальный гриб Uromyces fabae d By.
Спермогонии и эции формируются на нижней стороне листьев небольшими группами, часто концентрическими кругами. Эциоспоры шаровидные или эллиптические, размером 14-21 х 18-26 мкм, с мелкобородавчатой оболочкой.
Урединиопустулы размером до 1,5 мм, образуются с обеих сторон листа. Урединио-споры одноклеточные, шаровидные или яйцевидные, 17-26 х 21-30 мкм, желтоватые, с оболочкой, покрытой редкими шипиками.
Телиопустулы также формируются с обеих сторон листа. Телиоспоры 25-40 х 18-29 мкм, на желтоватой ножке, одноклеточные, округлые или яйцевидные, к основанию суженные, часто на вершине сосочкообразно вытянутые, с гладкой бурой оболочкой.
Гриб зимует на остатках растений в виде телиоспор, которые весной, прорастая, образуют базидии с базидиоспорами. От них и происходит первичное заражение растений.
При интенсивном развитии болезни недобор урожая зеленой массы и семян вики может достигать 20-30%.
Серая гниль
 Проявляется на пораженных листьях, стеблях, цветоносах и бобах в виде буровато-зеленых расплывчатых пятен, покрывающихся густым серым налетом. Позже в пораженных местах появляются черные склероции длиной 2-7 мм. Болезнь интенсивно развивается во второй половине вегетационного периода в условиях повышенной влажности воздуха. При наступлении сухой погоды пораженные места приобретают вид буроватых сухих язв.
Проявляется на пораженных листьях, стеблях, цветоносах и бобах в виде буровато-зеленых расплывчатых пятен, покрывающихся густым серым налетом. Позже в пораженных местах появляются черные склероции длиной 2-7 мм. Болезнь интенсивно развивается во второй половине вегетационного периода в условиях повышенной влажности воздуха. При наступлении сухой погоды пораженные места приобретают вид буроватых сухих язв.
Возбудитель болезни — многоядный несовершенный гриб Botrytis cinerea Fr. Он образует многочисленное конидиальное спороношение в виде серого налета. Конидиями гриб распространяется в период вегетации растений.
Сохраняется возбудитель в виде склероциев в почве и с семенами. Нередко грибница проникает в оболочку семян и тогда всхожесть их снижается.
С ерая гниль приводит к значительному уменьшению урожая зерна. Сортов, устойчивых к этой болезни, нет. Меньше поражаются растения сортов Рамонский 77 и Превосходный 240 (лущильный).
Темный аскохитоз
 Проявляется на листьях, стеблях и бобах в виде темно-коричневых пятен неправильной формы, размером 0,5-7 мм (большие пятна, как правило, зональные). Пикниды образуются на более крупных пятнах и разбросаны по всей поверхности. На пораженных стеблях часто образуются язвообразные углубления, а на всходах чернеет и загнивает корневая шейка, что иногда бывает причиной выпадения растений. На пораженных семенах хорошо заметны темные пятна.
Проявляется на листьях, стеблях и бобах в виде темно-коричневых пятен неправильной формы, размером 0,5-7 мм (большие пятна, как правило, зональные). Пикниды образуются на более крупных пятнах и разбросаны по всей поверхности. На пораженных стеблях часто образуются язвообразные углубления, а на всходах чернеет и загнивает корневая шейка, что иногда бывает причиной выпадения растений. На пораженных семенах хорошо заметны темные пятна.
Возбудителем темного аскохитоза является преимущественно гриб Ascochyta pinodes Jones. Кроме гороха, он поражает и другие зернобобовые культуры, но в меньшей степени. Гриб имеет бесполое и половое споро-ношения. При бесполом образуются пикниды и пикноспоры. Пикниды приплюснуто-шаровидные, темно-бурые, 165-180 мкм в диаметре. Пикноспоры бесцветные, цилиндрические, с закругленными или притупленными концами, размером 10-21 X 2,7- 6,1 мкм, с 1-3 перегородками.
Половое спороношение формируется обычно на засыхающих частях растений в виде мелких темно-коричневых, почти черных точек — псевдотециев с сумками и сумкоспорами. Сумки булавовидные или цилиндрические, сидячие или с очень короткой ножкой, размером 50-75 X 11,5-13,5 мкм. В каждой сумке обычно насчитывается по 8 бесцветных округлоэллиптических двух-клеточных сумкоспор размером 15-22 X 5- 8 мкм. В сумчатой стадии гриб имеет название Didymella pinodes Petr.
Пикноспоры и сумкоспоры возбудителя темного аскохитоза прорастают при относительной влажности воздуха выше 90%. Температурный оптимум для заражения растений в пределах 16-20°.
Болезни гречихи
- Печать
Аскохитоз
 Поражает листья и стебли, вследствие чего на них появляются большие округлые желтоватые пятна с темной каймой и нередко концентрической зональностью. В центре пятен в виде черных точек находится большое количество пикнид. Листья пораженных растений преждевременно усыхают и опадают.
Поражает листья и стебли, вследствие чего на них появляются большие округлые желтоватые пятна с темной каймой и нередко концентрической зональностью. В центре пятен в виде черных точек находится большое количество пикнид. Листья пораженных растений преждевременно усыхают и опадают.
Возбудитель болезни — несовершенный гриб Ascochyta fagopyri Bres. Его грибница межклеточная, пикниды шаровидные, 130-140 мкм в диаметре. Пикноспоры цилиндрические или эллиптические, бесцветные, двухклеточные, размером 16-18 х 6-7 мкм.
Распространяется гриб в период вегетации растений пикноспорами, а зимует на растительных остатках в виде грибницы и пикнид. Иногда пикниды могут находиться и на кожуре семени.
Недобор урожая гречихи от аскохитоза обычно не превышает 5-7%.
Бактериоз
 Проявляется обычно в период бутонизации и цветения. На листьях образуются сначала небольшие одиночные маслянистые темно-бурые округлые пятна до 5 мм в диаметре. Позже они сливаются, а пораженные листья сморщиваются и преждевременно засыхают.
Проявляется обычно в период бутонизации и цветения. На листьях образуются сначала небольшие одиночные маслянистые темно-бурые округлые пятна до 5 мм в диаметре. Позже они сливаются, а пораженные листья сморщиваются и преждевременно засыхают.
Болезнь вызывают бактерии Pseudomonas syringae van Hall. Это неспороносные с закругленными концами грамотрицательные палочки размером 0,3-0,5 х 1-3 мкм.
Сохраняются бактерии в неперегнивших остатках пораженных растений и в семенах. Могут распространяться ветром и дождем с кусочками пораженных тканей, а также насекомыми. При сильном поражении растений недобор зерна может достигать 8- 10%.
Ложная мучнистая роса (пероноспороз)
 Широко распространенное заболевание гороха в районах с достаточным увлажнением. Чаще проявляется в начале бутонизации растений. Поражаются все надземные органы, наблюдается местное и диффузное распространение болезни.
Широко распространенное заболевание гороха в районах с достаточным увлажнением. Чаще проявляется в начале бутонизации растений. Поражаются все надземные органы, наблюдается местное и диффузное распространение болезни.
При местном поражении с верхней стороны листьев появляются округлые беловатые или желтоватые пятна с нерезкими очертаниями. С нижней стороны листьев в местах пятен во влажную погоду образуется серовато-фиолетовый паутинистый налет. На стеблях и бобах возникают расплывчатые хлоротичные пятна, а затем и налет.
При диффузном поражении верхушки стеблей и листьев расположены близко друг к другу, вследствие чего растение напоминает головку цветной капусты. Серо-фиолетовый налет равномерно покрывает все надземные органы.
Возбудитель болезни — низший гриб Peronospora pisi Sydow. Его грибница межклеточная, конидиальное спороношение в виде налета образуется на поверхности пораженных растений, а ооспоры — в пораженных тканях.
Конидиеносцы коричневато-фиолетовые, дихотомически разветвленные, размером 250-450 X 6-9 мкм, выходят из устьиц по 1 -11 штук и часто образуют дернинки. На концах конидиеносцев формируются эллиптические бесцветные или желтоватые конидии размером 21-26 X 17-22 мкм.
Ооспоры шаровидные, диаметром 40-50 мкм, желтовато-коричневые, с толстой складчатой оболочкой.
Заболевание интенсивно развивается при высокой влажности и пониженных температурах. Источником инфекции являются остатки пораженных растений, в которых зимуют ооспоры. Иногда возбудитель сохраняется в виде грибницы в оболочке семян.
Пероноспороз очень вредоносен. Пораженные им растения отстают в росте, часто имеют карликовый вид, образуют щуплое зерно. В годы эпифитотии недобор урожая зерна может достигать 5-7 ц/га. Устойчивых сортов нет.
Мозаика
 Проявляется в виде желтой крапчатости и посветления вдоль жилок. Нередко междоузлия у пораженных растений укорочены, что придает ему вид карликовости. При повышении температуры воздуха до 30-35° внешние признаки заболевания часто маскируются, а при понижении до 20-25° проявляются вновь.
Проявляется в виде желтой крапчатости и посветления вдоль жилок. Нередко междоузлия у пораженных растений укорочены, что придает ему вид карликовости. При повышении температуры воздуха до 30-35° внешние признаки заболевания часто маскируются, а при понижении до 20-25° проявляются вновь.
Возбудитель болезни — вирус Cucumis virus I Smith. Передается инокуляцией сока, тлями и другими сосущими насекомыми. Носителями вируса могут быть также растения, в которых он сохраняется. С семенами вирус не передается. Меньше поражаются ранние посевы гречихи.
Филлостиктоз
 Проявляется на листьях в виде мелких, диаметром 2-4 мм, округлых беловатых пятен со светло-красной каймой. На пятнах расположены черные пикниды. Сильно пораженные листья преждевременно отмирают.
Проявляется на листьях в виде мелких, диаметром 2-4 мм, округлых беловатых пятен со светло-красной каймой. На пятнах расположены черные пикниды. Сильно пораженные листья преждевременно отмирают.
Возбудитель болезни — несовершенный гриб Phyllosticta polygonorum Sacc. Его пикниды шаровидные, диаметром до 170 мкм. Пикноспоры эллиптические или цилиндрические, бесцветные, одноклеточные, размером 5-8,4 х 2-2,8 мкм.
Сохраняется гриб в виде пикнид с пикноспорами на опавших листьях.
Болезни злаковых трав
- Печать
Белая пятнистость
 Поражает тимофеевку, лисохвост, ежу сборную, райграс и некоторые другие злаковые травы. На зеленых листьях с обеих сторон появляются светло-бурые удлиненные пятна с темно-пурпуровым ободком. В центре пятна, преимущественно с верхней стороны, образуются беловатые пушистые скопления конидиального спороношения возбудителя болезни — несовершенного гриба Mastigosporium album Riess.
Поражает тимофеевку, лисохвост, ежу сборную, райграс и некоторые другие злаковые травы. На зеленых листьях с обеих сторон появляются светло-бурые удлиненные пятна с темно-пурпуровым ободком. В центре пятна, преимущественно с верхней стороны, образуются беловатые пушистые скопления конидиального спороношения возбудителя болезни — несовершенного гриба Mastigosporium album Riess.
Конидиеносцы палочковидные, одноклеточные, бесцветные, длиной до 10 мкм. Конидии веретеновидные, бесцветные, с 3- 5 поперечными перегородками, размером 29-39 х 10-17 мкм (иногда 47-60 х 11-13 мкм), с одной-двумя ресничками на поверхности.
При помощи конидий гриб распространяется и заражает листья растений в период вегетации. При усыхании листьев в местах поражения формируются небольшие темные склероции, в виде которых гриб и зимует. Весной с прорастанием склероциев образуются новые конидии, заражающие растения. Особенно интенсивно болезнь распространяется во влажную погоду.
Преждевременное усыхание листьев приводит к уменьшению урожая зеленой массы и семян злаковых трав.
Головня
 Возбудителями ее являются различные базидиальные грибы порядка Ustilaginales, поражающие только определенные виды растений.
Возбудителями ее являются различные базидиальные грибы порядка Ustilaginales, поражающие только определенные виды растений.
На лисохвосте возбудителями головни являются Ustilago alopecurivora Liro и Urocystis alopecuri Frank. Первый образует на листьях и влагалищах узкие, почти черные полосы, вначале прикрытые эпидермисом. Через некоторое время эпидермис растрескивается, и из трещин распыляется черная споровая масса (телиоспоры). Часто пораженные листья растрескиваются и расщепляются в продольном направлении.
Телиоспоры шаровидные или эллиптические, размером 10-15 х 7-13 мкм, с желто-коричневой оболочкой, покрытой бородавкообразными выступами.
При поражении грибом Urocystis alopecuri на листьях и стеблях появляются серовато-черные продольные штрихи, просвечивающиеся через эпидермис, вследствие чего эту болезнь часто называют штриховатой листовой головней. Со временем пораженные ткани растрескиваются, и из них распыляются телиоспоры. Они собраны в неправильношаровидные темно-коричневые клубочки, состоящие из 1-3 центральных темных спор диаметром 7-12 мкм, с бородавчатой оболочкой, и нескольких бесцветных спор диаметром 4-10 мкм, расположенных по периферии.
На еже сборной подобную головню вызывают грибы Urocystis dactylidina M. Chochr. и Ustilago salvei Berk, et Br. У первого из них телиоспоры собраны в клубочки, состоящие из 1-4 темно-бурых яйцевидных спор размером 9-21 x 8-16 мкм и 5-11 бесцветных или темно-коричневых с гладкой оболочкой спор размером 7-14 х 3-5 мкм, расположенных по периферии. У второго телиоспоры не собраны в клубочки, шаровидные или эллиптические, размером 8-12 х 7-14 мкм, с желтовато-коричневой бородавчатой оболочкой.
На райграсе высоком часто обнаруживается листовая головня, вызываемая грибом Urocystis arrhenatheri M. Chochr. Телиоспоры его собраны в спорокучки, состоящие из 1-2 центральных шаровидных или яйцевидных спор размером 12,5-16 х 7-11 мкм, с желто-бурой оболочкой и нескольких периферических желтоватых спор диаметром 2-3,5 мкм.
В большинстве случаев заражение трав возбудителями головневых заболеваний происходит в почве во время прорастания семян. У пораженных растений не формируются генеративные органы, резко снижается урожай зеленой массы и семян. При сильном развитии болезни, когда поражено более 5% растений, сено нельзя использовать на корм скоту.
Серая пятнистость (овуляриоз)
Поражает ежу сборную, житняк, костер и некоторые другие злаковые травы. На листьях с верхней стороны появляются расплывчатые черноватые или охряно-бурые пятна, которые через некоторое время сливаются. С нижней стороны листьев образуется беловатый налет — конидиальное спороношение несовершенного гриба Ramularia jiusilla Unger (син. Ovularia pulchella Sacc). Конидиеносцы в пучках, чаще простые, слегка узловатые, размером 60-70 х 2-3,5 мкм. Конидии яйцевидные, размером 6-13 x 4,5-10 мкм. Распространяется гриб конидиями. Зимует на пораженных опавших листьях в виде грибницы, на котор ой весной образуется новое конидиальное спороношение.
ой весной образуется новое конидиальное спороношение.
Болезнь вызывает преждевременное отмирание листьев, что приводит к уменьшению урожая сена и семян трав.
Черная пятнистость
 Поражает тимофеевку, лисохвост, ежу сборную, житняк, райграс и некоторые другие злаковые травы. На листьях с верхней стороны появляются черные, блестящие, а в нижней — матовые овальные пятна длиной 0,5-1 мм. Нередко пятна сливаются и удлиняются до 20 мм. Вокруг них часто появляется светло-желтая кайма.
Поражает тимофеевку, лисохвост, ежу сборную, житняк, райграс и некоторые другие злаковые травы. На листьях с верхней стороны появляются черные, блестящие, а в нижней — матовые овальные пятна длиной 0,5-1 мм. Нередко пятна сливаются и удлиняются до 20 мм. Вокруг них часто появляется светло-желтая кайма.
Возбудитель болезни — сумчатый гриб Phyllachora graminis Fckl. Черные пятна на листьях являются плотным сплетением гиф грибницы (стромы). В строме образуются камеры перитециев, в которых формируются цилиндрические, размером 63-90 x 5,5-7 мкм, сумки на коротких ножках с парафизами. В каждой сумке располагаются в ряд цепочкой одноклеточные эллиптические желтоватые сумкоспоры размером 7-11 х 4,5-6 мкм.
Распространяется возбудитель болезни сумкоспорами, а зимует в виде перитециев на опавших листьях.
Болезнь интенсивно распространяется во влажную погоду и чаще обнаруживается очагами. Поражение растений приводит к снижению урожая сена и семян злаковых трав.
Чехловатость
Распространена почти на всех злаковых травах и характеризуется появлением преимущественно на верхней части стебля молочно-белого чехлика длиной около 4 см. Приблизительно через 3-4 недели чехлик уплотняется и приобретает золотисто-желтую окраску, а на его поверхности появляются черные точки. Чехлик является плотным сплетением грибницы — стромой возбудителя сумчатого гриба Epichloe typhina Tul. На строме формируется его беловатое конидиальное спороношение: короткие шилоподобные конидиеносцы размером 9-16 х 2,5-3 мкм и яйцевидные одноклеточные конидии размером 4-6X2-3 мкм. В конидиальной стадии гриб часто называют Sphacelia typhina Sacc.
Конидии, распространяясь, заражают молодые побеги злаковых трав.
С уплотнением грибницы в строме образуются перитеции, имеющие вид черных точек. Они грушевидные, размером 0,4-0,6 х 0,2-0,25 мм, с выводными отверстиями. В перитециях формируются сумки с сумкоспорами. Сумки цилиндрические, снизу суженные, размером 130-200 х 7-10 мкм, сумкоспоры бесцветные, нитевидные, одноклеточные, размером 80-160 х 1 — 1,5 мкм. Созревают они в конце лета, после выхода из сумок прорастают и образуют вторичные конидии, которые заражают растения через конус роста.
Гриб зимует в виде грибницы на пораженных растениях.
Вредоносность заболевания особенно велика в годы с влажным и прохладным летом. Пораженные растения не образуют генеративных органов и сильно отстают в росте. Болезнь может быть причиной недобора большого количества сем ян.
ян.
Болезни картофеля
- Печать
Альтернариоз
 Широко распространен в Белоруссии, западных и центральных районах европейской части России и особенно сильно — на Дальнем Востоке. Поражаются листья, стебли, а иногда и клубни картофеля. На листьях, обычно по краям долек, появляются мелкие темно-бурые пятна с густым бархатистым оливковым налетом. Сильно пораженные листья в сухую погоду скручиваются дольками кверху в виде лодочки.
Широко распространен в Белоруссии, западных и центральных районах европейской части России и особенно сильно — на Дальнем Востоке. Поражаются листья, стебли, а иногда и клубни картофеля. На листьях, обычно по краям долек, появляются мелкие темно-бурые пятна с густым бархатистым оливковым налетом. Сильно пораженные листья в сухую погоду скручиваются дольками кверху в виде лодочки.
Стебли и черешки листьев покрываются черными пятнами, но без заметной концентричности, как это бывает при макроспориозе. Поражение клубней встречается реже и характеризуется появлением округлых, слегка вдавленных пятен, которые могут покрываться черным налетом.
Возбудитель болезни — несовершенный гриб Alternaria solani Sor. Конидии его собраны в цепочки, бурые, обратнобулавовидные, с поперечными (4-8) и продольными (1-2) перегородками, размером 50- 80 X 11-12 мкм. Оптимальными условиями для прорастания конидий и заражения растений являются температура 22-26° и наличие капельной влаги в течение не менее двух часов.
Распространяется гриб конидиями, зимует на растительных остатках в виде грибницы и конидий. Иногда может сохраняться и на клубнях. Поражает многие виды семейства пасленовых.
Бугорчатая парша
 Проявляется через 4-5 месяцев после закладки картофеля на хранение. К весне признаки заболевания усиливаются. На больных клубнях образуются темные нерастрескивающиеся бугорки диаметром 3-4 мм с вдавленной бороздкой у основания. Расположены они одиночно или сливаются в кучки по три-пять штук в каждой.
Проявляется через 4-5 месяцев после закладки картофеля на хранение. К весне признаки заболевания усиливаются. На больных клубнях образуются темные нерастрескивающиеся бугорки диаметром 3-4 мм с вдавленной бороздкой у основания. Расположены они одиночно или сливаются в кучки по три-пять штук в каждой.
При совместном развитии бугорчатой и серебристой парши поверхностные ткани клубня темнеют, становятся неровными и шелушатся. Такой тип поражения иногда называют «черной кожей».
Возбудитель болезни — несовершенный гриб Oospora pustulans Owen et Wakef. Его грибница бесцветная, с перегородками, толщиной 2-2,5 мкм. Конидиеносцы многоклеточные, ветвящиеся, с цепочками конидий. Последние одноклеточные, овально цилиндрические, размером 6-12 X 2- 2,5 мкм. Интенсивность развития гриба усиливается при температуре 11-12°.
Клубни заражаются еще в поле через чечевички, глазки, механические повреждения кожуры, язвы парши. Заболевание сильнее проявляется на дерново-подзолистых песчаных и слабее на торфяных почвах.
Вертицеллезное увядание
 Внешним признаком является увядание растений.
Внешним признаком является увядание растений.
Возбудитель — гриб Verticilliurn alboatrum Reinke et Berth.). Особенно большой вред они причиняют на юге и юго-востоке страны. Проявляются в начале цветения растений картофеля. При вертициллезном увядании прежде всего теряют тургор и желтеют края отдельных долек листа. Затем на листьях появляются светло-бурые пятна с ярко-желтой каймой. В сухую погоду листья засыхают и опадают, во влажную — повисают вдоль стебля.
На черешках и главной жилке увядших листьев может появляться серовато-грязный налет, стебли также отмирают, но стоят до уборки. На поперечном срезе стебля и корня можно обнаружить потемнение сосудистых пучков. На поверхности кусочков пораженных стеблей, помещенных во влажную камеру, через несколько дней появляется сероватый налет, состоящий из конидиеносцев и конидий гриба. Конидиеносцы прямостоячие, у основания темноокрашенные, до 700 мкм длиной, мутовчато разветвленные, несущие одноклеточные яйцевидные или продолговатые бесцветные конидии размером 5-12 X 2-3 мкм.
Кроме конидий, гриб образует хламидоспоры, которые зимуют на остатках растений и в почве. Грибница может проникать в глазки клубней, вследствие чего во время хранения они загнивают и превращаются в серую пылящую массу. На месте глазков образуются впадинки. Гриб может вызывать также увядание томата, баклажана, перца, люпина, хмеля и других растений. Источниками инфекции являются зараженные клубни картофеля, пораженные остатки растений и зараженная хламидоспорами почва.
Заболевание нарушает водоснабжение надземных частей растений, что приводит к их увяданию и отмиранию. Особенно большой вред причиняет картофелю в засушливых юго-восточных районах.
При фузариозном увядании листья, начиная с верхних, теряют тургор, приобретают бледно-зеленую окраску. Верхушка стебля становится антоциановой и поникает. За ночь тургор на некоторое время восстанавливается, но затем листья полностью желтеют, скручиваются, и все растение засыхает. У пораженных растений разрушается подземный стебель, буреют боковые корни и столоны.
Гриб образует макро- и микроконидии, хламидоспоры, а иногда и склероции. Макроконидии формируются на воздушной грибнице, реже в спородохиях. По форме они могут быть веретено-серповидные, эллиптические или изогнутые. Микроконидии одно-, двухклеточные, хламидоспоры гладкие или шероховатые, одно- или двухклеточные, бесцветные, диаметром до 12 мкм.
В растение гриб проникает через корневые волоски, а затем по ксилеме продвигается в стебель и закупоривает сосуды, вызывая их интоксикацию.
Готика
 Больные растения вытянуты, листья их мелкие, морщинистые, со слабо скрученными дольками параллельно средней жилке, темно-зеленой или фиолетовой окраски, от стебля отходят под более острым углом, чем у здоровых растений. Клубни больных растений в большинстве случаев веретеновидные, многоглазковые, неправильной формы, в засушливых условиях у некоторых сортов покрываются крупными некротичными пятнами и трещинами. Возбудитель болезни — вироид, существующий в виде нуклеиновой кислоты без белкового компонента. Передается заболевание с клубнями и семенами картофеля.
Больные растения вытянуты, листья их мелкие, морщинистые, со слабо скрученными дольками параллельно средней жилке, темно-зеленой или фиолетовой окраски, от стебля отходят под более острым углом, чем у здоровых растений. Клубни больных растений в большинстве случаев веретеновидные, многоглазковые, неправильной формы, в засушливых условиях у некоторых сортов покрываются крупными некротичными пятнами и трещинами. Возбудитель болезни — вироид, существующий в виде нуклеиновой кислоты без белкового компонента. Передается заболевание с клубнями и семенами картофеля.
Аукуба-мозаика. На листьях нижнего, а у некоторых сортов и среднего яруса образуются ярко-желтые округлые пятна. Иногда появляются некрозы коленхимы стеблей и листьев, что вызывает опадение нижних, а позже и средних листьев. У некоторых сортов на клубнях после двух-трех месяцев хранения образуются некрозы в виде коричневых пятен неправильной формы в сердцевине и на кожуре.
Возбудитель болезни — Solanum virus 9 Smith — передается клубнями, инокуляцией и тлями.
Дуплистость клубней
 Проявляется в виде полостей или пустот нередко звездчатой формы. Стенки полости ровные, сухие и покрыты слоем опробковевшей ткани. Пораженные клубни более водянистые.
Проявляется в виде полостей или пустот нередко звездчатой формы. Стенки полости ровные, сухие и покрыты слоем опробковевшей ткани. Пораженные клубни более водянистые.
Дуплистость возникает вследствие чрезмерно быстрого роста клубней. Особенно часто это наблюдается у крупноклубневых сортов во влажную теплую погоду на хорошо унавоженных полях.
Железистая (ржавая пятнистость) клубней
 Внешне не проявляется. Однако в мякоти клубней образуются отмершие, опробковевшие участки от 0,1 до 2 см в поперечнике, ржаво-коричневой окраски различных оттенков — от янтарного до красно-бурого. Пятнистость чаще располагается по окружности клубня, вблизи камбиального слоя. При сильном поражении отмершие участки сливаются и проникают в более глубокие слои. В кипящей воде пораженная мякоть клубня не разваривается.
Внешне не проявляется. Однако в мякоти клубней образуются отмершие, опробковевшие участки от 0,1 до 2 см в поперечнике, ржаво-коричневой окраски различных оттенков — от янтарного до красно-бурого. Пятнистость чаще располагается по окружности клубня, вблизи камбиального слоя. При сильном поражении отмершие участки сливаются и проникают в более глубокие слои. В кипящей воде пораженная мякоть клубня не разваривается.
В период хранения клубней заболевание не распространяется.
Причиной железистой пятнистости являются неблагоприятные почвенные условия в период формирования и роста клубней (сухость и высокая температура почвы, недостаток элементов минерального питания, особенно калия).
Закручивание листьев
 Проявляется в виде мозаичности и закручивания верхних листьев. Общий хлороз растений, кожистость и хрупкость листьев не наблюдаются, чем это заболевание отличается от скручивания листьев.
Проявляется в виде мозаичности и закручивания верхних листьев. Общий хлороз растений, кожистость и хрупкость листьев не наблюдаются, чем это заболевание отличается от скручивания листьев.
Возбудитель болезни — вирус М — распространяется с зараженным посадочным материалом и переносится тлями.
Картофельная гниль (фитофтороз)
 Встречается во многих районах страны. Поражаются листья, стебли и клубни, особенно в начале цветения картофеля. На листьях, начиная с нижних, а также на отдельных участках стебля появляются небольшие, но быстро увеличивающиеся темно-бурые пятна. Листья увядают, чернеют и засыхают, а во влажную погоду загнивают. На нижней стороне листа вокруг пятен на границе с зеленой здоровой тканью в сырую погоду или при утренней росе образуется беловатый паутинистый налет.
Встречается во многих районах страны. Поражаются листья, стебли и клубни, особенно в начале цветения картофеля. На листьях, начиная с нижних, а также на отдельных участках стебля появляются небольшие, но быстро увеличивающиеся темно-бурые пятна. Листья увядают, чернеют и засыхают, а во влажную погоду загнивают. На нижней стороне листа вокруг пятен на границе с зеленой здоровой тканью в сырую погоду или при утренней росе образуется беловатый паутинистый налет.
На клубнях появляются резко очерченные сероватые, а затем бурые вдавленные твердые пятна различной величины. При разрезе клубня в его периферической части видна ржавчина мякоти, распространяющаяся к центру в виде язычков.
Возбудитель болезни — низший гриб Phytophthora infestans d By. Грибница его несептирована, распространяется в межклетниках тканей. На поверхность листа через устьица выступают в виде налета зооспорангиеносцы с зооспорангиями. Зооспорангиеносцы слабо разветвлены — имеют от одной до четырех основных ветвей и несколько боковых с утолщениями в местах образования зооспорангиев. Последние одноклеточные, овальные, размером 25-33 X 15-20 мкм.
В период вегетации растений гриб распространяется зооспорангиями. Прорастают они в капельножидкой влаге, образуя зооспоры или инфекционный росток, который внедряется в ткани растений.
Продолжительность инкубационного периода заболевания в зависимости от температуры от 3 до 16 дней. Минимальной температурой, при которой может происходить развитие гриба в растении, является 1,3, а максимальной — около 30°.
Клубни заражаются либо от зооспорангиев, которые при обильных дождях могут с листьев попадать в почву, либо во время уборки урожая, когда они соприкасаются с поверхностным слоем почвы или с пораженной ботвой. Механические повреждения клубней способствуют проникновению гриба.
Во время хранения картофеля заболевание не распространяется, но в местах фитофторозных поражений часто поселяются многие микроорганизмы, нередко вызывающие гниение клубней в хранилищах.
В природных условиях на территории нашей страны ооспоры гриба не образуются. Зимует гриб в форме грибницы в клубнях. Первые признаки заболевания в поле наблюдаются на ростках картофеля.
При сильном поражении ботвы недобор урожая составляет 70% и более.
Относительно устойчивы к фитофторозу сорта картофеля Житомирянка, Мета, Мостовский, Темп, Сулев, Кандидат, Белорусский крахмалистый, Столовый 19, Львовянка, Комсомолец, Домодедовский, Садко и др.
Крапчатая мозаика
 Проявляется на листьях в виде светло-зеленой крапчатости, а на некоторых сортах картофеля в виде черных некротических пятен. Возбудители заболевания — вирусы X и S, которые распространяются насекомыми. Резерваторами их могут быть дурман, томат, белена и другие растения семейства пасленовых.
Проявляется на листьях в виде светло-зеленой крапчатости, а на некоторых сортах картофеля в виде черных некротических пятен. Возбудители заболевания — вирусы X и S, которые распространяются насекомыми. Резерваторами их могут быть дурман, томат, белена и другие растения семейства пасленовых.
Мокрая бактериальная гниль клубней
 Развивается в поле на переувлажненных участках, но наиболее сильно проявляется в период хранения картофеля. Пораженные клубни становятся темно-бурыми, мягкими и мокрыми, их мякоть превращается в кашицеобразную или слизистую тягучую массу, издающую неприятный запах.
Развивается в поле на переувлажненных участках, но наиболее сильно проявляется в период хранения картофеля. Пораженные клубни становятся темно-бурыми, мягкими и мокрыми, их мякоть превращается в кашицеобразную или слизистую тягучую массу, издающую неприятный запах.
Возбудителями мокрой гнили являются различные полусапрофитные бактерии, преимущественно Erwinia carotovora Holl. var. atroseptica Tennison и Pseudomonas xanthochlora Stapp. Они проникают в клубни через повреждения, а также поселяются на участках, пораженных фитофторозом, паршой и другими болезнями.
Мокрая гниль быстро развивается при повышенной влажности воздуха и резких колебаниях температуры в местах хранения.
Морщинистая мозаика
 Вызывает сильное вздутие листьев между жилками и скручивание долек листа вниз. В первый год заражения растений заболевание обычно проявляется слабо, но в потомстве (на второй-третий годы) оно вызывает отставание растений в росте, укорачивание междоузлий и образование мелких курчавых хлоротичных листьев. У некоторых сортов наблюдается полегание стеблей. Больные растения не цветут и заканчивают вегетацию на три-четыре недели раньше здоровых. Часто листья и стебли становятся хрупкими и преждевременно отмирают.
Вызывает сильное вздутие листьев между жилками и скручивание долек листа вниз. В первый год заражения растений заболевание обычно проявляется слабо, но в потомстве (на второй-третий годы) оно вызывает отставание растений в росте, укорачивание междоузлий и образование мелких курчавых хлоротичных листьев. У некоторых сортов наблюдается полегание стеблей. Больные растения не цветут и заканчивают вегетацию на три-четыре недели раньше здоровых. Часто листья и стебли становятся хрупкими и преждевременно отмирают.
Морщинистая мозаика относится к заболеваниям со смешанным типом инфекции. Ее вызывает вирус Solanum virus 2 Smith в комбинации с вирусами X, А, S, а иногда К и М.
Вирусы передаются с клубнями, распространяются тлями и другими сосущими насекомыми.
Обыкновенная парша
 Распространена повсеместно. На поверхности клубней, преимущественно на чечевичках, появляются неглубокие неправильно-округлой формы язвочки от нескольких миллиметров до 1 см и более в диаметре. Сливаясь, они часто образуют сплошную корку. Поражаются также столоны и корни.
Распространена повсеместно. На поверхности клубней, преимущественно на чечевичках, появляются неглубокие неправильно-округлой формы язвочки от нескольких миллиметров до 1 см и более в диаметре. Сливаясь, они часто образуют сплошную корку. Поражаются также столоны и корни.
Возбудителями болезни являются актиномицеты, чаще Streptomyces scabies Waks. et Heur. (син. Actinomyces scabies Gus-sow). На основных нитях их грибницы вырастают воздушные гифы с винтообразно закрученными спороносцами, на которых формируются мелкие цилиндрические продолговатые споры размером 1,2-0,8 X 0,8-1 мкм. Оптимальная для развития патогена температура 25-27°.
При выкопке картофеля из влажной почвы в глубине язвочек парши часто можно видеть сероватый, зеленоватый или другого цвета налет лучистых грибов, который на открытом воздухе быстро исчезает.
Актиномицеты, вызывающие обыкновенную паршу, обитают в почве на различных органических остатках. Сильнее поражают клубни на легких и суглинистых почвах со слабощелочной реакцией.
Меньше поражаются сорта Богатырь, Вита, Гатчинский, Детскосельский, Олев, Петровский, Столовый 19, Янтарный и др.
Полосчатая мозаика
 Характеризуется появлением некротических темных полосочек, точек и пятен на жилках и в уголках между ними (угловатая пятнистость). Листья становятся хрупкими, темнеют, отмирают и опадают или остаются висеть на тонких высохших черешках под острым углом к главному стеблю. Некрозы также обнаруживаются на черешках листьев и стеблях.
Характеризуется появлением некротических темных полосочек, точек и пятен на жилках и в уголках между ними (угловатая пятнистость). Листья становятся хрупкими, темнеют, отмирают и опадают или остаются висеть на тонких высохших черешках под острым углом к главному стеблю. Некрозы также обнаруживаются на черешках листьев и стеблях.
Возбудитель этой болезни — Solanum virus 2 Smith (вирус Y). Передается он тлями и механическим путем вместе с инфицированным соком, а зимует в клубнях картофеля. В последнее время также наблюдается поражение картофеля вирусом, вызывающим хлоротичность, крапчатость и бугристость листьев.
Нередко пораженные растения полегают и преждевременно отмирают.
Порошистая парша
 Чаще встречается на торфяных почвах. Поражает клубни, корни, столоны и участки стеблей, находящиеся в почве.
Чаще встречается на торфяных почвах. Поражает клубни, корни, столоны и участки стеблей, находящиеся в почве.
На клубнях образуются язвы с обрывками перидермы и кожицы, напоминающие форму звездочек. На дне их заметна порошкообразная бурая споровая масса.
На корнях, столонах и стеблях появляются наросты различной величины и формы, расположенные одиночно или группами. Вначале они белые, позднее темнеют и распадаются.
Возбудитель болезни — низший гриб Spongospora subterranea (Wallr.) John-son. Это внутриклеточный паразит, который в вегетирующем состоянии представляет собой бесформенный комочек протоплазмы в виде амебоида, способный к самостоятельному передвижению. Достигая подземных органов растений, он проникает в его клетки, где постепенно разрастается в плазмодий. Затем в плазмодии происходит деление ядра, и вокруг каждого вторичного ядра обособляется участок протоплазмы. На таких новообразованиях появляются одноклеточные мелкие многоугольные споры 2-4 мкм в диаметре. Обычно они склеены в полые неправильной формы шарообразные кучки 42-50 мкм в диаметре, которыми бывают заполнены язвы на клубнях. Спорокучки могут также попадать в почву, где сохраняются до пяти лет.
Оптимальными условиями для развития заболевания являются повышенная влажность почвы (около 70% от полной влагоемкости) и температура 12-18°. От начала заражения до проявления первых признаков заболевания проходит около 12, а до полного формирования спор — около 29 дней.
Сортов картофеля, высокоустойчивых к этой болезни, нет.
Почернение сердцевины клубней
 В мякоти клубней появляются расплывчатые желтовато-серые участки, постепенно приобретающие желтовато-бурую, а затем черную окраску. Сначала пораженные участки небольшие, но по мере развития заболевания они могут распространяться на большую часть клубня. При сильном поражении из побуревшей мякоти выделяются капельки желтоватой мутной жидкости.
В мякоти клубней появляются расплывчатые желтовато-серые участки, постепенно приобретающие желтовато-бурую, а затем черную окраску. Сначала пораженные участки небольшие, но по мере развития заболевания они могут распространяться на большую часть клубня. При сильном поражении из побуревшей мякоти выделяются капельки желтоватой мутной жидкости.
Пораженные клубни теряют товарные и вкусовые качества. При варке они еще сильнее чернеют, но не затвердевают, как при железистой пятнистости.
Сердцевина темнеет вследствие нарушения ферментативных процессов в клубне при неблагоприятных условиях транспортировки и хранения: повышении температуры, недостатке кислорода, накоплении углекислоты, травмировании клубней.
Ранняя сухая пятнистость (макроспориоз)
 Проявляется на листьях за 15-20 дней до цветения картофеля в виде крупных темно-бурых округлых или угловатых пятен с концентрическими кругами и слабым черным налетом с нижней стороны листа. Ткань в местах пятен высыхает и продырявливается. На стеблях появляются темно-бурые концентрические пятна с черным налетом.
Проявляется на листьях за 15-20 дней до цветения картофеля в виде крупных темно-бурых округлых или угловатых пятен с концентрическими кругами и слабым черным налетом с нижней стороны листа. Ткань в местах пятен высыхает и продырявливается. На стеблях появляются темно-бурые концентрические пятна с черным налетом.
Возбудитель болезни — несовершенный гриб Macrosporium solani ЕU. et Mart. На пораженных тканях образуются его булавовидные дымчато-серые конидии размером 100-200 X 12-20 мкм. Конидия разделена поперечными (7-10) и продольными (1-3) перегородками и имеет длинный придаток.
Распространяются конидии в период вегетации растений ветром и каплями дождя. При их прорастании образуется инфекционный росток, который проникает в ткани растений через устьица. Грибница межклеточная. Гриб выделяет токсин, вызывающий пожелтение и отмирание тканей.
Заболевание особенно сильно развивается в жаркую погоду с обильными дождями и росами. Инкубационный период его длится от трех до восьми дней. Конидии могут формироваться при температуре от 15 до 34,5°. Болезнь усиливается при недостатке калия в почве.
Сохраняется возбудитель на перезимовавших в почве послеуборочных остатках, с которых в следующем году попадает на здоровые растения.
Кроме картофеля, поражает белену, белладонну, томаты и другие растения семейства пасленовых.
Вредоносность болезни особенно велика при поражении вегетативной массы, вследствие чего уменьшается ассимиляционная поверхность, а в конечном счете — урожай клубней и их качество.
Повышенную устойчивость имеют сорта Филатовский, Бородянский, Огонек, Гатчинский, Зарево и др.
Северный столбур
 Характеризуется замедлением роста кустов картофеля, чрезмерным ветвлением, мелколистностью и позеленением цветков, но увядание растений не наблюдается.
Характеризуется замедлением роста кустов картофеля, чрезмерным ветвлением, мелколистностью и позеленением цветков, но увядание растений не наблюдается.
Заболевание передается зараженными клубнями. Насекомые-переносчики не обнаружены.
Серебристая парша
 При поражении клубней в почве их кожура покрывается многочисленными пятнами темно-бурого цвета диаметром от 1 до 6 мм с легко стирающимся налетом — спороношением возбудителя болезни.
При поражении клубней в почве их кожура покрывается многочисленными пятнами темно-бурого цвета диаметром от 1 до 6 мм с легко стирающимся налетом — спороношением возбудителя болезни.
На хранящихся клубнях в местах пятен спороношения нет, но под кожурой обнаруживается тонкая белая грибница, которая со временем темнеет и покрывается клубочками склероциев.
В местах поражения ткань пробковеет, а кожура отслаивается от мякоти. Пятна имеют коричневый цвет с серебристым оттенком, который хорошо заметен при смачивании клубней водой.
Возбудитель болезни — несовершенный гриб Helminthosporium solani Dur. et Mont. (син. Spondylocladium atrovirens Harz. et Sacc.). В местах поражения на молодых клубнях при повышенных температурах (23-25°) из склероциальных клубочков образуются конидиеносцы с конидиями. Конидиеносцы прямые, темно-оливковые, длиной 150-500, толщиной 4,8 мкм. Конидии обратнобулавовидные, черно-оливковые, размером 36-61 X 8-11 мкм, с четырьмя-шестью перегородками и заостренными концами, расположены в несколько ярусов мутовками по две-четыре штуки.
При сплошном поражении верхней части клубня почки отмирают и всходы не образуются. Источниками инфекции являются пораженные клубни и почва, где гриб сохраняется на растительных остатках.
Серебристая парша часто встречается на клубнях, пораженных бугорчатой паршой. Устойчивых к болезни сортов нет.
Стеблевая нематода
 Ditylenchus destructor Thorne. Поражает клубни и столоны картофеля, корни хмеля и сирени, луковицы лилейных культур и корневища мяты. Пораженные растения отстают в росте, их стебли утолщены и укорочены, листья мелкие, волнистые и хлоротичные, со скрученными внутрь волнистыми краями. До начала клубнеобразования личинки нематод поражают также корневую шейку и стебли картофеля.
Ditylenchus destructor Thorne. Поражает клубни и столоны картофеля, корни хмеля и сирени, луковицы лилейных культур и корневища мяты. Пораженные растения отстают в росте, их стебли утолщены и укорочены, листья мелкие, волнистые и хлоротичные, со скрученными внутрь волнистыми краями. До начала клубнеобразования личинки нематод поражают также корневую шейку и стебли картофеля.
Однако признаки поражения надземных органов не всегда типичны. Наиболее характерно поражение клубней. Вначале на молодых клубнях в местах поражения кожура буреет и приобретает вид жирного пятна. Затем она становится бархатистой или шелковистой, а окраска ее меняется до серовато-черной с разными оттенками в зависимости от сорта картофеля. Позже в этих местах появляются волнистые морщины, кожура растрескивается и, ссыхаясь, сморщивается. При хранении такие клубни полностью сморщиваются и загнивают вследствие развития на них гнилостных грибов и бактерий.
Взрослые особи нематоды червеобразные. Самки размером 0,72-1,44 X 0,022-0,032, а самцы 0,75-1,3 X 0,020-0,025 мм. Зимуют в почве в стадии яиц, хотя в состояние анабиоза могут переходить во всех стадиях своего развития. Личинки отрождаются и поражают картофель при температуре от 5 до 34° и влажности выше 40%. При 20-24° развитие одного поколения длится 20-26 дней. За вегетационный период может развиваться 6-9 поколений.
Недобор урожая от стеблевой нематоды иногда достигают значительных размеров — 10-20%.
Сухая гниль клубней
 Проявляется на клубнях во время хранения. На их поверхности появляются серовато-бурые
Проявляется на клубнях во время хранения. На их поверхности появляются серовато-бурые
или матовые, несколько вдавленные в мякоть пятна различной формы. Мякоть клубня становится бурой, трухлявой и сухой. Позже пятна увеличиваются, ткани в этих местах сморщиваются и на их поверхности появляются небольшие выпуклые серовато-белые, реже желтоватые или розоватые подушечки. В сухом хранилище клубень, пораженный гнилью, постепенно высыхает, а его кожица сморщивается в виде складок. При хранении картофеля в сыром помещении (влажность воздуха выше 90%) гниль становится мокрой, но не превращается в слизистую с неприятным запахом массу.
Возбудители болезни — грибы из рода Fusarium Link, чаще F. solani App. et Wr. Гифы их, переплетаясь, образуют плотные подушечки различной окраски, на поверхности которых формируются короткие конидиеносцы с серповидными конидиями. Последние различаются окраской и количеством перегородок в зависимости от вида возбудителя.
Болезнь распространяется конидиями и грибницей. Оптимальными условиями их развития являются температура 17-25°, относительная влажность воздуха 70% и достаточная аэрация.
Проникая в мякоть клубня через повреждения, грибы разрушают стенки клеток, и протоплазму. Нетронутыми остаются лишь крахмальные зерна.
Сохраняются возбудители болезни на клубнях, растительных остатках и в почве. При интенсивном развитии заболевания потери урожая могут достигать 30% и более.
Трахеобактериозное увядание
 Вызывают бактерии Pseudomonas solanacearum Berg., Corynebacterium sepedonicum Skapt et Burkh. Первые чаще встречаются на юге и Дальнем Востоке. Пораженные ими растения быстро увядают, листья их желтеют, сморщиваются, черешки Листьев и стебли поникают.
Вызывают бактерии Pseudomonas solanacearum Berg., Corynebacterium sepedonicum Skapt et Burkh. Первые чаще встречаются на юге и Дальнем Востоке. Пораженные ими растения быстро увядают, листья их желтеют, сморщиваются, черешки Листьев и стебли поникают.
Поражение растений бактерией С. sepedonicum обычно более распространено в северной и центральной зонах страны. Признаки заболевания обнаруживаются в конце цветения картофеля. Листья постепенно желтеют, дольки их скручиваются вдоль главной жилки, стебли поникают и засыхают. Особенно заметны признаки заболевания в сухое, жаркое лето.
На разрезе больного клубня, а иногда и стебля пораженный участок сосудистой системы имеет лимонно-желтый цвет и однородную маслянистую консистенцию. При надавливании из пораженного места выступает светло-желтая тягучая масса. Часто такое поражение называют кольцевой гнилью клубней.
Бактерии могут вызывать также ямчатую гниль клубней, которая обнаруживается лишь в конце марта. При очистке клубней от кожуры заметны округлые с булавочную головку пятна гниющей мякоти, имеющие желтоватую или кремовую окраску. Вокруг этих пятен образуется более прозрачная, но еще твердая мякоть. Позже ямчатая гниль углубляется, расширяется: кожица клубня над гнилью лопается и на поверхности его образуется вдавленная ямка. При повышенной температуре (18-20°) развитие ямчатой гнили усиливается.
Относительно устойчивы к этому заболеванию сорта картофеля Ульяновский, Смачный и др.
Фомоз (пуговичная гниль)
 Заболевание распространено на Дальнем Востоке, в Сибири, Поволжье и других районах возделывания картофеля. Поражаются клубни и стебли. На клубнях образуются темные твердые вдавленные пятна, достигающие 2,5-5 см в диаметре. Часто эти пятна имеют вид пуговицы. Поэтому такое поражение клубней иногда называют пуговичной болезнью. На поверхности пятен образуются многочисленные черные пикниды, выступающие через кожицу клубня. Вначале поражения ткань клубня имеет светло-коричневую окраску, а позже темнеет, сморщивается и становится темно-серой или черной. Часто пораженная ткань загнивает и образуются пустоты, внутри которых развивается рыхлая войлочная грибница возбудителя. В отличие от сухой гнили, в местах поражения на клубнях отсутствуют концентрические круги сморщенной ткани и конидиальное спороношение в виде подушечек.
Заболевание распространено на Дальнем Востоке, в Сибири, Поволжье и других районах возделывания картофеля. Поражаются клубни и стебли. На клубнях образуются темные твердые вдавленные пятна, достигающие 2,5-5 см в диаметре. Часто эти пятна имеют вид пуговицы. Поэтому такое поражение клубней иногда называют пуговичной болезнью. На поверхности пятен образуются многочисленные черные пикниды, выступающие через кожицу клубня. Вначале поражения ткань клубня имеет светло-коричневую окраску, а позже темнеет, сморщивается и становится темно-серой или черной. Часто пораженная ткань загнивает и образуются пустоты, внутри которых развивается рыхлая войлочная грибница возбудителя. В отличие от сухой гнили, в местах поражения на клубнях отсутствуют концентрические круги сморщенной ткани и конидиальное спороношение в виде подушечек.
На стеблях болезнь проявляется во время цветения картофеля. У основания черешков листьев по всей поверхности стебля образуются удлиненные расплывчатые пятна, на которых формируются многочисленные, сначала светлые, а позже темно-коричневые или черные пикниды. Пятна часто охватывают до 2/3 окружности стебля, а по длине достигают 8 см. На поздних сортах картофеля наряду с образованием пятен на стеблях возникают удлиненноовальные язвы глубиной 1 -1,5 мм и длиной до 4 см.
Вокруг язв ткань стебля отмирает и приобретает коричневый оттенок.
Возбудитель болезни — несовершенный гриб Phoma exigua Desm. (син. Ph. tuberosa M., R. et Sch.). В его пикнидах образуются светлые одноклеточные пикно-споры размером 4-6 X 2-3,5 мкм. Во влажную погоду пикниды на стеблях раскрываются, и споры с капельками дождя или ветром разносятся на посевах, вызывая новое заражение стеблей. Вместе с дождевой водой споры попадают через почву на вновь образовавшиеся клубни, вызывая их заражение.
Гриб проникает в клубни через чечевички, глазки и поврежденную кожицу.
В период хранения клубней фомоз часто развивается одновременно с фузариозной гнилью, вызывая большие потери продукции. Первичным источником инфекции являются пораженные клубни. Есть данные о том, что в почве возбудитель может сохраняться до трех лет.
Фузариозное увядание
 Внешним признаком является увядание растений.
Внешним признаком является увядание растений.
Возбудитель — гриб Fusarium oxysporum Schl.). Особенно большой вред они причиняют на юге и юго-востоке страны. Проявляются в начале цветения растений картофеля. При вертициллезном увядании прежде всего теряют тургор и желтеют края отдельных долек листа. Затем на листьях появляются светло-бурые пятна с ярко-желтой каймой. В сухую погоду листья засыхают и опадают, во влажную — повисают вдоль стебля.
На черешках и главной жилке увядших листьев может появляться серовато-грязный налет, стебли также отмирают, но стоят до уборки. На поперечном срезе стебля и корня можно обнаружить потемнение сосудистых пучков. На поверхности кусочков пораженных стеблей, помещенных во влажную камеру, через несколько дней появляется сероватый налет, состоящий из конидиеносцев и конидий гриба. Конидиеносцы прямостоячие, у основания темноокрашенные, до 700 мкм длиной, мутовчато разветвленные, несущие одноклеточные яйцевидные или продолговатые бесцветные конидии размером 5-12 X 2-3 мкм.
Кроме конидий, гриб образует хламидоспоры, которые зимуют на остатках растений и в почве. Грибница может проникать в глазки клубней, вследствие чего во время хранения они загнивают и превращаются в серую пылящую массу. На месте глазков образуются впадинки. Гриб может вызывать также увядание томата, баклажана, перца, люпина, хмеля и других растений. Источниками инфекции являются зараженные клубни картофеля, пораженные остатки растений и зараженная хламидоспорами почва.
Заболевание нарушает водоснабжение надземных частей растений, что приводит к их увяданию и отмиранию. Особенно большой вред причиняет картофелю в засушливых юго-восточных районах.
При фузариозном увядании листья, начиная с верхних, теряют тургор, приобретают бледно-зеленую окраску. Верхушка стебля становится антоциановой и поникает. За ночь тургор на некоторое время восстанавливается, но затем листья полностью желтеют, скручиваются, и все растение засыхает. У пораженных растений разрушается подземный стебель, буреют боковые корни и столоны.
Гриб образует макро- и микроконидии, хламидоспоры, а иногда и склероции. Макроконидии формируются на воздушной грибнице, реже в спородохиях. По форме они могут быть веретено-серповидные, эллиптические или изогнутые. Микроконидии одно-, двухклеточные, хламидоспоры гладкие или шероховатые, одно- или двухклеточные, бесцветные, диаметром до 12 мкм.
В растение гриб проникает через корневые волоски, а затем по ксилеме продвигается в стебель и закупоривает сосуды, вызывая их интоксикацию.
Черная ножка
 Проявляется в виде загнивания нижней части стебля молодых растений. Рост больных растений замедляется и часто прекращается. Нижние листья становятся кожистыми, ломкими, с загнутыми вверх краями, а верхние скручиваются и остаются мелкими. Черешки листьев располагаются под более острым углом к главному стеблю. В зависимости от сорта и погодных условий загнивающие участки стебля могут приобретать различную окраску: светло- и темно-зеленую, коричневую, темно-бурую, желтую, фиолетовую.
Проявляется в виде загнивания нижней части стебля молодых растений. Рост больных растений замедляется и часто прекращается. Нижние листья становятся кожистыми, ломкими, с загнутыми вверх краями, а верхние скручиваются и остаются мелкими. Черешки листьев располагаются под более острым углом к главному стеблю. В зависимости от сорта и погодных условий загнивающие участки стебля могут приобретать различную окраску: светло- и темно-зеленую, коричневую, темно-бурую, желтую, фиолетовую.
На более взрослых растениях во влажную, прохладную погоду болезнь проявляется в верхней части стебля в виде сплошного ослизнения молодых тканей, которые приобретают темно-зеленую окраску. На черешках и дольках листьев появляются коричневые пятна и полоски. На поперечном срезе больного стебля видно почернение сосудов.
В годы с небольшими осадками и жарким летом возбудители болезни могут переходить в клубни нового урожая без внешних признаков поражения (скрытая форма заболевания). При раннем развитии черной ножки растения не образуют клубней, а при более позднем клубни хотя и образуются, но многие из них поражаются внутри черной гнилью, всегда начинающейся со столонной части.
Вызывают заболевание различные виды почвенных бактерий, главным образом Pectobacterium phytophthorum (Appel) Waldee (син. Erwinia phytophthora Berg. et all). В клубни бактерии проникают через столоны, чечевички и различные повреждения.
Источниками инфекции могут быть пораженные клубни и ботва, остающиеся в поле и не сгнивающие до весны. В период вегетации заболевание может распространяться насекомыми. Наибольший вред черная ножка причиняет в условиях повышенной влажности на тяжелых по механическому составу почвах.
Парша картофеля. Паршой называют различные поверхностные поражения клубней, вызываемые грибами. Известно пять видов парши: обыкновенная, черная, пыльная, серебристая и бугорчатая.
Черная парша
 Распространена в Белоруссии и полесской зоне Украины, на Дальнем Востоке, в нечерноземных областях России. Поражаются клубни, стебли, столоны и корни взрослых растений. На поверхности клубней образуются твердые черные, неправильной формы коростинки (склероции) различной величины, напоминающие комочки приставшей почвы. На ростках и корнях появляются несколько вдавленные пятна бурой окраски и язвы диаметром до 1 см и более. Иногда вместо них образуется буроватая штриховатость. Пораженный участок отмирает.
Распространена в Белоруссии и полесской зоне Украины, на Дальнем Востоке, в нечерноземных областях России. Поражаются клубни, стебли, столоны и корни взрослых растений. На поверхности клубней образуются твердые черные, неправильной формы коростинки (склероции) различной величины, напоминающие комочки приставшей почвы. На ростках и корнях появляются несколько вдавленные пятна бурой окраски и язвы диаметром до 1 см и более. Иногда вместо них образуется буроватая штриховатость. Пораженный участок отмирает.
Возбудитель болезни — базидиальный гриб Hypochnus solani Pr. et Del. В базидиальной стадии во влажную теплую погоду он поражает стебли картофеля, вследствие чего на них образуется грязно-белый войлочный налет, легко снимающийся в виде пленки. Такое поражение называют белой ножкой. Однако в большинстве случаев гриб развивает грибницу и склероции (коростинки на клубнях), поэтому его нередко называют Rhizoctonia solani Kuehn. и относят к несовершенным грибам, а причиняемое им поражение называют ризоктониозом. Развивается гриб при высокой влажности и температуре от 9 до 27° (оптимум 15-21°). Зимует он в виде склероциев на клубнях картофеля и в почве. После посадки больных клубней из прорастающих склероциев образуется грибница, которая проникает в развивающиеся ростки и вызывает их загнивание и гибель. Гриб поражает также свеклу, капусту, морковь, табак и многие другие растения. Менее поражаются сорта Садко, Гатчинский, Олев, Смена и др.
Южный столбур
 Характеризуется окаймлением долек верхушечных листьев, измельчением отрастающих, укорачиванием междоузлий и разрастанием пазушных листьев. Затем на 7-10-й день наблюдается частичное отмирание корневой системы и увядание растений. На клубнях пораженных растений образуются нитевидные бесцветные ростки без корней.
Характеризуется окаймлением долек верхушечных листьев, измельчением отрастающих, укорачиванием междоузлий и разрастанием пазушных листьев. Затем на 7-10-й день наблюдается частичное отмирание корневой системы и увядание растений. На клубнях пораженных растений образуются нитевидные бесцветные ростки без корней.
В период вегетации болезнь может передаваться от больных растений к здоровым цикадками.
Источник https://biomolecula.ru/articles/pochemu-boleiut-rasteniia-i-kak-oni-boriutsia-s-infektsiei-na-primere-fitopatogennykh-bakterii
Источник https://rosselhoscenter.com/index.php/2014-02-28-11-39-42/2011-11-16-12-58-47/bolezni
Источник
Источник